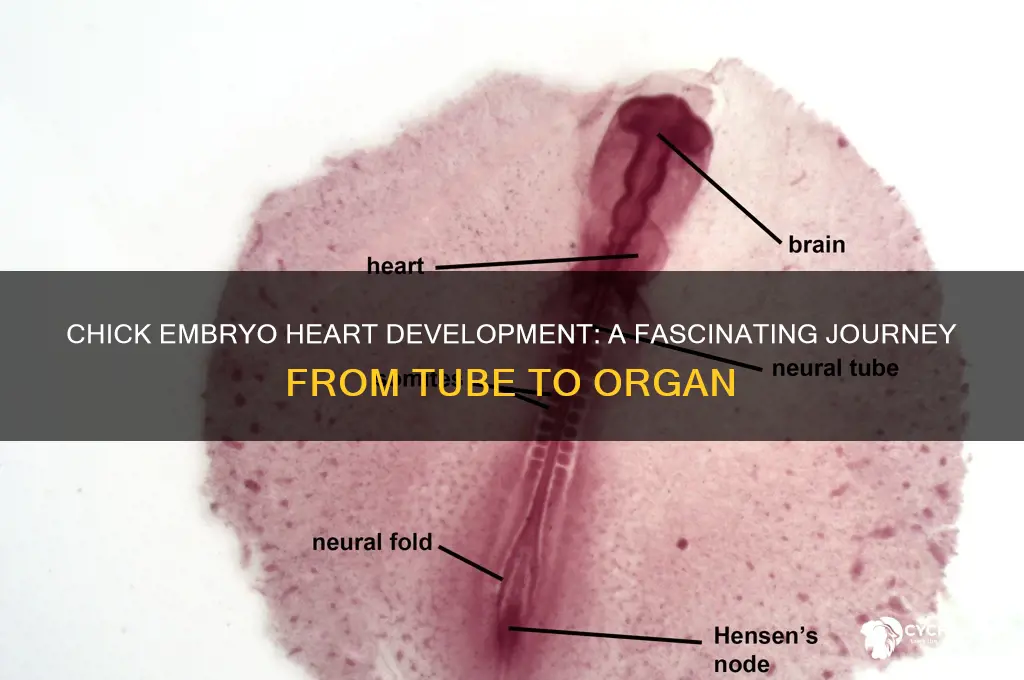
The development of the heart in a chick embryo is a fascinating and intricate process that begins early in embryogenesis. Within the first few days of incubation, the embryonic cells differentiate and migrate to form the primordial heart tube, a critical structure that lays the foundation for the cardiovascular system. This process, known as cardiogenesis, involves the coordinated expression of specific genes and signaling pathways that guide the formation, looping, and septation of the heart tube. As the embryo develops, the heart undergoes a series of morphological changes, transforming from a simple tube into a four-chambered organ capable of pumping blood efficiently. Understanding these developmental stages not only sheds light on the complexity of organogenesis but also provides valuable insights into congenital heart defects and potential therapeutic strategies.
| Characteristics | Values |
|---|---|
| Initiation of Heart Development | Begins around 24-30 hours after incubation (HH stage 4-5) with the formation of the splanchnic mesodermal layer adjacent to the endoderm. |
| Primitive Streak Formation | Mesoderm migrates from the primitive streak, forming the cardiogenic mesoderm, which will give rise to the heart tube. |
| Cardiogenic Mesoderm Specification | Induced by signals from the endoderm (e.g., BMP, Wnt, FGF, and Nodal) to form the primary heart field (PHF) and secondary heart field (SHF). |
| Heart Tube Formation | Occurs by HH stage 10 (36-40 hours). The PHF forms the initial heart tube through lateral migration and fusion of mesodermal cells. |
| Looping Morphogenesis | Begins around HH stage 12 (48-52 hours). The heart tube undergoes rightward looping, establishing the asymmetry necessary for chamber formation. |
| Chamber Differentiation | By HH stage 18 (60-72 hours), the heart tube differentiates into distinct chambers: atrium, ventricle, and outflow tract, driven by SHF contributions. |
| Septation | Atrial and ventricular septa form to separate the chambers into left and right sides, ensuring proper blood flow. |
| Coronary Vasculature Development | Begins around HH stage 22 (72-80 hours) with the formation of coronary vessels from the proepicardial organ. |
| Electrical Conduction System | Develops concurrently with chamber formation, establishing the sinoatrial node, atrioventricular node, and bundle of His. |
| Hematopoiesis | The heart tube also serves as a site for early hematopoiesis, producing blood cells before the liver and bone marrow take over. |
| Molecular Regulation | Key genes involved include Nkx2.5, Gata4, Tbx5, and Hand1/2, which regulate cardiomyocyte differentiation and morphogenesis. |
| Blood Flow Establishment | Functional blood flow begins around HH stage 14 (52-56 hours), facilitating nutrient and gas exchange in the developing embryo. |
| Maturation | The heart continues to mature throughout embryonic development, increasing in size and complexity to support post-hatching life. |
Explore related products






What You'll Learn
![]()
Stage-specific cardiac progenitor formation
The development of the heart in a chick embryo is a highly orchestrated process, involving the precise formation and differentiation of cardiac progenitor cells at specific stages. Stage-specific cardiac progenitor formation begins during gastrulation, around Hamburger-Hamilton (HH) stage 3 to 4, when the primitive streak forms and mesodermal cells are specified. These mesodermal progenitors, located in the anterior lateral plate mesoderm (ALPM), are fated to give rise to cardiac tissue. Key signaling pathways, such as BMP (Bone Morphogenetic Protein), Wnt, and FGF (Fibroblast Growth Factor), play critical roles in patterning and inducing cardiac progenitors. The ALPM cells express markers like *Mesp1* and *Isl1*, which are essential for their commitment to the cardiac lineage. This early stage sets the foundation for subsequent heart tube formation and is crucial for proper cardiac morphogenesis.
By HH stage 6 to 8, cardiac progenitors in the ALPM begin to migrate toward the midline, driven by chemotactic signals and changes in cell adhesion. This migration is a critical step in stage-specific cardiac progenitor formation, as it ensures the proper alignment of cells for heart tube assembly. During this stage, the progenitors further differentiate into distinct populations: the first heart field (FHF) and the second heart field (SHF). FHF progenitors contribute primarily to the left ventricle and parts of the atria, while SHF progenitors form the right ventricle, outflow tract, and additional atrial tissue. The SHF remains proliferative and continues to provide cells for heart growth throughout later stages of development.
At HH stage 10 to 12, the cardiac progenitors converge at the midline to form the cardiac crescent, marking the beginning of heart tube formation. This stage is characterized by the activation of cardiac-specific genes such as *Nkx2.5*, *Gata4*, and *Tbx5*, which drive the differentiation of progenitors into cardiomyocytes and other cardiac cell types. The epithelial-to-mesenchymal transition (EMT) is also initiated here, allowing cells to reorganize and form a linear heart tube. Proper coordination of these processes is essential for the structural integrity of the developing heart, and disruptions at this stage can lead to congenital heart defects.
Between HH stage 13 to 18, the heart tube undergoes looping, a process where it folds and twists to establish the asymmetry necessary for chamber formation. Stage-specific cardiac progenitor formation continues as SHF progenitors contribute to the elongation and growth of the heart tube. The outflow tract and atria expand, and the primordia of the future chambers become distinct. Signaling molecules like FGF10 and retinoic acid regulate SHF proliferation and deployment, ensuring the heart grows proportionally. This stage highlights the dynamic nature of cardiac progenitor specification, as cells continue to differentiate and integrate into the developing heart structure.
Finally, by HH stage 20 and beyond, chamber differentiation and septation occur, completing the formation of a four-chambered heart. While the focus shifts to maturation and functional integration, the earlier stage-specific cardiac progenitor formation remains the cornerstone of this process. The precise temporal and spatial regulation of progenitor specification, migration, and differentiation ensures the heart develops with the correct anatomy and functionality. Understanding these stages provides critical insights into both normal cardiac development and the mechanisms underlying congenital heart diseases.
Minimizing the Impact: Strategies for Chicken Barn Effects
You may want to see also
Explore related products






![]()
Endocardial and myocardial differentiation
The development of the heart in a chick embryo is a complex and highly coordinated process, involving the differentiation of various cell types, including endocardial and myocardial cells. Endocardial and myocardial differentiation is a critical step in heart development, as it establishes the inner lining of the heart (endocardium) and the muscular wall (myocardium), which are essential for proper heart function. This process begins during the early stages of embryonic development, specifically during the splanchnic mesodermal stage, when the cardiogenic mesoderm gives rise to the precursor cells that will eventually form the heart.
Endocardial differentiation initiates when the splanchnic mesoderm cells migrate to the midline and form the cardiogenic plate. As the cardiogenic plate folds and fuses to create the cardiac tube, a subset of cells within the tube begins to express markers characteristic of endocardial cells, such as Tie2 and NFATC1. These cells line the inner surface of the cardiac tube, forming the primitive endocardium. The differentiation of endocardial cells is regulated by signaling pathways, including BMP, Wnt, and Notch, which interact to specify the endocardial fate and promote the formation of a continuous endothelial layer. This layer is crucial for the subsequent development of the cardiac valves and the coronary vessels.
Myocardial differentiation occurs concurrently with endocardial differentiation but involves a distinct population of cells. Myocardial precursor cells, derived from the cardiogenic mesoderm, migrate to form the myocardium surrounding the endocardial tube. These cells undergo a mesenchymal-to-epithelial transition (MET), aligning and compacting to form the muscular walls of the heart. Key transcription factors such as Nkx2.5, GATA4, and MEF2C drive myocardial differentiation by activating cardiac-specific gene programs, including those for sarcomeric proteins and contractile machinery. The interaction between the endocardium and myocardium is essential, as signals from the endocardium, such as Neuregulin1, are critical for myocardial trabeculation and wall thickening.
The differentiation of endocardial and myocardial cells is tightly regulated by both intrinsic and extrinsic factors. Intrinsically, genetic programs control cell fate decisions, while extrinsically, signals from neighboring tissues and the extracellular matrix guide morphogenesis. For instance, the epicardium, which forms later in development, secretes factors like retinoic acid and FGFs that influence myocardial proliferation and maturation. Additionally, blood flow through the developing heart provides mechanical cues that further refine myocardial structure and function. Disruptions in these processes can lead to congenital heart defects, underscoring the importance of precise endocardial and myocardial differentiation.
In summary, endocardial and myocardial differentiation in the chick embryo is a multifaceted process involving distinct cellular programs, signaling pathways, and tissue interactions. The endocardium forms the inner lining of the heart, while the myocardium develops into the muscular walls, both of which are essential for cardiac function. Understanding these differentiation processes provides insights into normal heart development and the pathogenesis of cardiac abnormalities, offering potential targets for therapeutic intervention in congenital heart diseases.
White Chicken Chili: Spices to Warm Your Soul
You may want to see also
Explore related products






![]()
Role of signaling pathways (e.g., BMP, Wnt)
The development of the heart in a chick embryo is a complex and highly coordinated process that relies on precise signaling pathways to ensure proper formation and function. Among these, the Bone Morphogenetic Protein (BMP) and Wnt signaling pathways play critical roles in cardiac morphogenesis. BMP signaling is essential during the early stages of heart development, particularly in the formation of the cardiac mesoderm. BMP signals, primarily BMP2 and BMP4, are secreted by surrounding tissues and act on the lateral plate mesoderm to induce cardiac fate. These signals activate downstream effectors such as Smad proteins, which translocate to the nucleus and regulate the expression of cardiac transcription factors like Nkx2.5 and GATA4. This process is crucial for the specification of cardiogenic mesoderm, the precursor tissue that will give rise to the heart tube.
The Wnt signaling pathway is another key regulator of heart development in chick embryos, functioning both in the early specification of cardiac mesoderm and in later stages of cardiac morphogenesis. Canonical Wnt signaling, mediated by β-catenin, is active in the anterior lateral plate mesoderm and promotes the differentiation of cardiac progenitor cells. Wnt signals interact with BMP pathways to fine-tune the balance between cardiac and extracardiac fates, ensuring that the correct number and type of cells are allocated to the developing heart. Additionally, Wnt signaling is involved in the epithelial-to-mesenchymal transition (EMT) during cardiac cushion formation, a critical step in the development of heart valves and septa. Dysregulation of Wnt signaling can lead to congenital heart defects, highlighting its importance in this process.
The interplay between BMP and Wnt pathways is particularly noteworthy during the formation of the linear heart tube. BMP signaling helps maintain the anterior-posterior patterning of the heart by regulating the expression of homeobox genes like Msx1 and Msx2, which are essential for proper heart tube assembly. Simultaneously, Wnt signaling ensures the correct alignment and fusion of the bilateral heart fields. This coordination is achieved through feedback mechanisms where BMP signals modulate Wnt activity and vice versa, creating a robust network that guides cardiac morphogenesis. For example, BMP inhibitors like Noggin and Chordin are regulated by Wnt signals, ensuring that BMP activity is spatially and temporally restricted to allow for proper heart tube formation.
In later stages of heart development, both BMP and Wnt pathways continue to play vital roles in cardiac looping and chamber differentiation. BMP signaling is involved in the asymmetric growth of the heart tube, which is essential for the looping process that establishes the future atrial and ventricular chambers. Wnt signaling, on the other hand, regulates the proliferation and differentiation of cardiomyocytes, ensuring that the heart grows in size while maintaining its structural integrity. The precise regulation of these pathways is critical for the formation of a functional four-chambered heart, and disruptions can lead to severe cardiac abnormalities.
In summary, the BMP and Wnt signaling pathways are indispensable for the development of the heart in chick embryos. They act in concert to regulate cardiac mesoderm specification, heart tube formation, cardiac looping, and chamber differentiation. Through intricate crosstalk and feedback mechanisms, these pathways ensure the precise spatial and temporal control required for the complex process of cardiogenesis. Understanding their roles not only provides insights into normal heart development but also sheds light on the molecular basis of congenital heart defects, offering potential targets for therapeutic intervention.
Leaving Raw Chicken Out Overnight: Safe or Not?
You may want to see also
Explore related products






![]()
Looping morphogenesis in heart tube
The process of heart development in chick embryos is a fascinating journey, and looping morphogenesis plays a crucial role in shaping the initial heart tube into a functional organ. This intricate transformation occurs during the early stages of embryonic growth, setting the foundation for the circulatory system. Here's an in-depth look at this remarkable process:
Looping morphogenesis is a critical event in the transformation of the straight heart tube into a more complex structure. It involves the bending and folding of the tube, creating a looped configuration. This process is not merely a simple bend but a highly coordinated series of cellular movements and shape changes. As the heart tube forms from the fusion of the paired anterior heart fields, it initially lacks the characteristic shape of a mature heart. The looping phase is essential for establishing the asymmetry and specific morphology required for proper heart function. During this stage, the heart tube undergoes rightward and dextral looping, which is a complex 3D maneuver. The rightward looping brings the future ventricles to the right side, while dextral looping twists the tube, positioning the outflow tract anteriorly and the inflow tract posteriorly.
The mechanism driving this morphogenesis is a combination of cellular and molecular processes. Cells in the heart tube actively migrate and rearrange, contributing to the bending and twisting motions. This cellular rearrangement is guided by various signaling pathways and genetic factors. For instance, the left-right signaling pathway plays a pivotal role in initiating the looping process, ensuring that the heart develops with the correct lateral orientation. Additionally, the planar cell polarity pathway influences cell shape changes and coordinated movement, facilitating the intricate folding of the heart tube. These molecular signals create a dynamic environment, instructing cells to move and reshape, ultimately forming the looped structure.
As looping progresses, the heart tube transforms into a more recognizable heart shape. The initial loop establishes the primitive ventricle and atrium, with the outflow tract and inflow tract taking their respective positions. This process is not just about creating a curved shape but also involves the differentiation of specific heart regions. The bending and twisting motions bring different parts of the heart tube into proximity, allowing for the subsequent formation of septa and valves. The precision of looping morphogenesis is vital, as any deviations can lead to congenital heart defects.
In the chick embryo, this entire process occurs within a relatively short developmental window, showcasing the efficiency and complexity of embryonic growth. The study of looping morphogenesis provides valuable insights into the understanding and potential treatment of heart-related birth defects. By unraveling the molecular and cellular mechanisms, researchers can contribute to the broader field of developmental biology and regenerative medicine. This intricate dance of cells during looping morphogenesis is a testament to the remarkable precision of embryonic development.
Ligaments in Chicken Feet: Their Function and Role
You may want to see also
Explore related products





![]()
Septation and chamber formation process
The septation and chamber formation process in the developing chick embryo heart is a complex and highly coordinated sequence of events that transforms the initial tubular heart into a four-chambered structure. This process begins around embryonic day 3.5 (HH stage 16-18) and continues through day 6. Initially, the heart tube consists of a single lumen lined by myocardium and endocardium. Septation involves the formation of partitions (septa) that divide this tube into distinct chambers, ensuring proper segregation of oxygenated and deoxygenated blood. The process is driven by signaling pathways, cellular differentiation, and tissue remodeling, primarily involving the endocardial cushion tissue.
The first critical step in septation is the development of the endocardial cushions, which are swellings composed of mesenchymal cells derived from the endocardium. These cushions form at the atrioventricular (AV) canal and the outflow tract (OFT) regions. The AV cushions will eventually fuse to form the AV septum, separating the left and right atria from the ventricles. Simultaneously, the OFT cushions contribute to the formation of the aorticopulmonary septum, which divides the outflow tract into the aorta and pulmonary artery. Transforming growth factor-β (TGF-β) and bone morphogenetic protein (BMP) signaling pathways play a pivotal role in endocardial cushion formation and mesenchymal cell differentiation.
As the cushions develop, the heart tube begins to loop and align along the embryonic body axis, a process known as cardiac looping. This looping is essential for the spatial arrangement of the future chambers. By embryonic day 4, the heart has undergone rightward looping, positioning the ventricle to the left of the atrium. This alignment sets the stage for the subsequent formation of the interventricular septum (IVS), which divides the left and right ventricles. The IVS forms through the fusion of two components: the muscular IVS, derived from myocardium, and the membranous IVS, derived from the endocardial cushions.
Chamber specification and maturation occur concurrently with septation. The atria and ventricles become morphologically distinct due to differential growth and gene expression patterns. For instance, *Nkx2.5* and *GATA4* transcription factors are critical for ventricular identity, while *TBX5* is essential for atrial development. The trabeculation process, where myocardial cells proliferate and form intricate networks within the ventricular walls, also begins during this stage, increasing the surface area for efficient blood flow.
Finally, the completion of septation and chamber formation ensures the establishment of a functional four-chambered heart by embryonic day 6. Proper alignment and fusion of the septa are crucial to prevent congenital heart defects, such as septal defects or misalignment of the great arteries. The entire process is tightly regulated by genetic and epigenetic factors, highlighting the precision required in embryonic heart development. Understanding these mechanisms provides valuable insights into both normal cardiac morphogenesis and the etiology of congenital heart diseases.
Chicken Tenders Portion Sizes at TGI Fridays
You may want to see also
Frequently asked questions
The heart begins to form at approximately 1.5 to 2 days of incubation, during the gastrulation stage, when the splanchnic mesoderm migrates to form the cardiogenic mesoderm.
The heart tube forms through the fusion of paired cardiogenic mesoderm cells along the embryonic midline, creating a single, straight tube that will later loop and differentiate into distinct heart regions.
Heart looping is a critical morphogenetic event occurring around 2.5 days of incubation, where the heart tube bends and twists to establish the asymmetry necessary for the formation of atria, ventricles, and outflow tracts.
Blood flow begins around 3 days of incubation and is essential for heart development, as it stimulates the remodeling of the heart tube, promotes chamber differentiation, and ensures proper alignment of cardiac structures.
Key molecular signals include BMP (Bone Morphogenetic Protein), FGF (Fibroblast Growth Factor), and Wnt pathways, which regulate cell differentiation, migration, and patterning during cardiogenesis.































