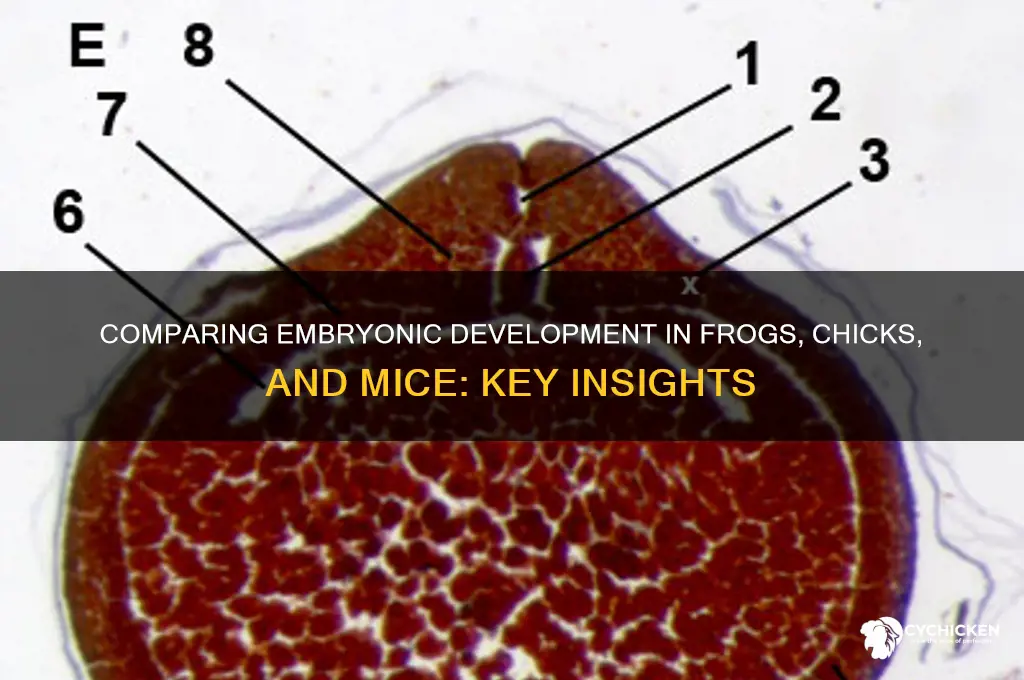
The study of embryology in frogs, chicks, and mice provides a fascinating comparative framework for understanding the diverse developmental processes across species. Frogs, as amphibians, undergo external fertilization and exhibit rapid, visible developmental stages, making them ideal for studying early embryonic patterning and organogenesis. Chick embryos, with their large size and accessibility, offer valuable insights into gastrulation, neural tube formation, and limb development, often serving as a bridge between simpler and more complex vertebrate models. Mice, as mammals, provide a critical perspective on internal fertilization, placental development, and the intricate molecular mechanisms driving organ differentiation, while also being a cornerstone of genetic research. Together, these models highlight both conserved and divergent pathways in vertebrate development, offering a comprehensive understanding of the fundamental principles governing life’s earliest stages.
Explore related products




$249.99 $159


What You'll Learn
- Gastrulation Process: Formation of germ layers (ectoderm, mesoderm, endoderm) in frog, chick, and mouse embryos
- Neural Tube Development: Early nervous system formation and differences across species during embryogenesis
- Somite Formation: Segmentation of paraxial mesoderm into somites in frog, chick, and mouse
- Organogenesis Timing: Comparative timeline of organ development in frog, chick, and mouse embryos
- Yolk Utilization: Role of yolk in nutrient supply and embryonic growth across species
![]()
Gastrulation Process: Formation of germ layers (ectoderm, mesoderm, endoderm) in frog, chick, and mouse embryos
The gastrulation process is a critical phase in embryonic development where the three primary germ layers—ectoderm, mesoderm, and endoderm—are formed, laying the foundation for all future tissues and organs. In frog embryos, gastrulation begins with the formation of a blastopore, which serves as the opening to the archenteron (primitive gut). Cells from the dorsal lip of the blastopore migrate inward in a process called involution, forming the mesoderm and endoderm layers. The ectoderm remains as the outermost layer. This process is highly coordinated and involves extensive cell movements, including convergence and extension, which narrow and elongate the embryo. The mesoderm splits into two layers: the dorsal mesoderm, which contributes to structures like the notochord, and the ventral mesoderm, which forms organs such as the heart and blood vessels. The endoderm lines the archenteron and gives rise to internal organs like the digestive and respiratory systems.
In chick embryos, gastrulation is similarly marked by the formation of a primitive streak, a structure analogous to the blastopore in frogs. Cells from the primitive streak migrate to form the mesoderm and endoderm layers, while the ectoderm remains on the surface. The process involves epithelial-mesenchymal transition (EMT), where cells lose their epithelial characteristics and become motile mesenchymal cells. The chick embryo’s gastrulation is more complex due to the larger size and greater cell number, requiring precise regulation of cell movements. The mesoderm divides into paraxial, intermediate, and lateral plate mesoderm, each contributing to specific structures such as somites, kidneys, and the circulatory system. The endoderm forms the gut tube, which later differentiates into organs like the liver and pancreas.
Mouse embryos also undergo gastrulation via the formation of a primitive streak, but the process is more rapid and tightly regulated. Cells from the proximal end of the streak migrate to form the mesoderm and endoderm, while the ectoderm persists as the outer layer. Mouse gastrulation involves the node, a structure that acts as the organizer and directs cell movements. The mesoderm is further organized into axial, paraxial, and lateral plate mesoderm, contributing to the notochord, somites, and internal organs, respectively. The endoderm forms the yolk sac and gut tube, which develop into the digestive and respiratory systems. Unlike frogs and chicks, mouse embryos rely heavily on signaling pathways like Wnt and BMP for patterning and cell fate determination during gastrulation.
Comparatively, while the gastrulation process in frogs, chicks, and mice shares the common goal of forming the three germ layers, the mechanisms and structures involved differ. Frogs rely on a blastopore and involution, chicks use a primitive streak and EMT, and mice employ a primitive streak and node-based signaling. Despite these differences, all three organisms exhibit coordinated cell movements and signaling pathways to ensure proper layer formation. Understanding these processes provides insights into the conserved and divergent aspects of vertebrate development, highlighting the complexity and precision of embryogenesis across species.
Bae Catches Cheating: What Now?
You may want to see also
Explore related products






![]()
Neural Tube Development: Early nervous system formation and differences across species during embryogenesis
The development of the neural tube, a critical structure that gives rise to the central nervous system (CNS), is a highly conserved yet species-specific process during embryogenesis. In frogs, chicks, and mice, neural tube formation begins with gastrulation, where the ectodermal layer is induced to form the neural plate. In frogs, this process is influenced by signals from the organizer region (equivalent to the Spemann organizer), which secretes factors like BMP antagonists to pattern the neuroectoderm. The neural plate then folds along the midline, a process driven by coordinated cell shape changes and convergent extension, ultimately fusing to form the neural tube. This rapid development in frogs is adapted for their aquatic embryonic environment, with the neural tube becoming evident within the first few days of development.
In chicks, neural tube development is similarly initiated by signals from the node (the avian equivalent of the organizer), which establishes the anterior-posterior axis. The neural plate in chicks undergoes extensive convergent extension movements, facilitated by planar cell polarity (PCP) pathways, to narrow and elongate the tissue. Unlike frogs, chick embryos develop within a shell, and their neural tube formation is synchronized with the establishment of extra-embryonic membranes. The process is slightly slower compared to frogs but is highly regulated to ensure proper closure, as defects can lead to neural tube defects (NTDs), a common embryological issue in avian species.
Mice exhibit a more complex and protracted neural tube development due to their mammalian physiology. Gastrulation in mice involves the formation of the primitive streak, which organizes the embryo into three germ layers. Neural induction is regulated by a combination of signals, including BMP, Wnt, and FGF antagonists, which pattern the ectoderm into neural fate. The neural plate folds and fuses along the entire length of the embryo, a process that must be precisely coordinated to prevent NTDs such as spina bifida. Unlike frogs and chicks, mice undergo secondary neurulation in the spinal region, adding another layer of complexity to their neural tube development.
Despite these species-specific adaptations, the molecular mechanisms underlying neural tube development are conserved. For instance, the role of Sonic Hedgehog (Shh) in patterning the ventral neural tube is essential across all three species, though its expression domains and timing differ. Similarly, Notch signaling regulates neurogenesis in all species, but the timing and spatial distribution of Notch activity vary, reflecting the distinct developmental timelines and environmental constraints of each organism.
Key differences in neural tube development across these species highlight evolutionary adaptations to their respective environments and reproductive strategies. Frogs, with their external aquatic development, prioritize rapid neural tube closure to protect the embryo. Chicks, developing within a shell, balance rapid growth with the need for proper closure to avoid NTDs. Mice, as mammals, have a prolonged and intricate process that supports the development of a complex CNS. Understanding these differences provides insights into the evolutionary plasticity of neural tube development while underscoring the conserved molecular pathways that drive early nervous system formation.
Chicken Patty Mystery: What's in the Chicken Big Mac?
You may want to see also
Explore related products






![]()
Somite Formation: Segmentation of paraxial mesoderm into somites in frog, chick, and mouse
Somite formation is a critical process in the embryological development of vertebrates, including frogs, chicks, and mice, marking the segmentation of the paraxial mesoderm into distinct blocks called somites. These somites give rise to essential structures such as the vertebral column, ribs, dermis, and skeletal muscle. The process begins with the differentiation of the paraxial mesoderm, which is positioned adjacent to the neural tube and notochord. In all three organisms, the notochord plays a pivotal role in inducing the overlying mesoderm to form somites through signaling molecules like Sonic Hedgehog (Shh). This initial step sets the stage for the rhythmic and sequential segmentation of the paraxial mesoderm into somites, a process regulated by the segmentation clock, a molecular oscillator involving genes such as *Hes7* in mice and their orthologs in frogs and chicks.
In frogs, somite formation occurs during the early stages of embryogenesis, shortly after gastrulation. The paraxial mesoderm is organized into pairs of somites on either side of the notochord and neural tube. The process is highly synchronized, with new somites forming at regular intervals from the anterior to the posterior end of the embryo. The segmentation clock in frogs involves oscillations of genes like *Her1* and *Her7*, which are homologous to the mouse *Hes7*. The precise timing and patterning of somite formation in frogs are influenced by the interplay between the notochord, neural tube, and lateral plate mesoderm, with signals such as Wnt and FGF pathways playing crucial roles in maintaining the segmentation clock and boundary formation.
Chicks exhibit a similar but more complex somite formation process due to their amniote characteristics. The paraxial mesoderm in chicks is initially unsegmented and forms a structure called the presomitic mesoderm (PSM). As development progresses, the PSM undergoes rhythmic segmentation into somites, with the process regulated by the segmentation clock involving *Hes7* oscillations. The chick embryo’s somite formation is also influenced by gradients of retinoic acid and FGF signaling, which help establish the anterior-posterior axis and maintain the periodicity of somite segmentation. The epithelialization of newly formed somites in chicks is particularly notable, as it involves the rapid rearrangement of cells into a polarized epithelium, a process driven by cadherin-mediated cell adhesion.
In mice, somite formation is a well-studied model for understanding the molecular mechanisms of segmentation. Similar to chicks, the mouse embryo develops a PSM, which is segmented into somites in a rostral-to-caudal direction. The segmentation clock in mice is driven by oscillations of *Hes7*, with each oscillation corresponding to the formation of one somite pair. The periodicity of *Hes7* expression is regulated by a negative feedback loop involving its protein product. Additionally, the Notch signaling pathway is essential for maintaining the oscillations and ensuring proper somite boundary formation. Mutations in genes involved in this process, such as *Dll1* or *Lfng*, lead to defects in somite segmentation, highlighting their critical role in mouse embryogenesis.
Comparatively, while the fundamental mechanisms of somite formation are conserved across frogs, chicks, and mice, there are species-specific differences in the molecular players and the timing of segmentation. For instance, the segmentation clock genes and signaling pathways exhibit variations in their expression patterns and interactions. Despite these differences, the rhythmic segmentation of the paraxial mesoderm into somites remains a hallmark of vertebrate development, underscoring the evolutionary conservation of this process. Understanding somite formation in these model organisms provides valuable insights into the broader principles of embryology and the developmental origins of musculoskeletal structures.
Easy Tips to Securely Keep Your Chicken Wrap Closed Every Time
You may want to see also
Explore related products

![]()
Organogenesis Timing: Comparative timeline of organ development in frog, chick, and mouse embryos
Organogenesis, the process by which organs form and develop, varies significantly in timing and sequence across different vertebrate species. When comparing frog, chick, and mouse embryos, the differences in organogenesis timing reflect their distinct developmental strategies and evolutionary adaptations. Frogs, as amphibians, develop rapidly in aquatic environments, while chicks (birds) and mice (mammals) have longer gestation periods and more complex organ systems. Understanding these timelines provides insights into the conserved and divergent mechanisms of embryogenesis.
In frog embryos, organogenesis begins remarkably early, often within the first few days of development. The neural tube, a precursor to the central nervous system, forms by stage 12 (approximately 24 hours post-fertilization). By stage 18 (around 48 hours), the heart tube is visible and begins to beat, marking the onset of cardiovascular development. Limb buds appear by stage 22, and major organs such as the liver, pancreas, and kidneys start to differentiate by stage 25. This rapid pace of organogenesis is driven by the need for quick adaptation to the aquatic environment, with tadpoles becoming free-swimming within a week.
Chick embryos exhibit a more extended timeline for organogenesis, reflecting their amniotic development and the complexity of avian systems. The neural tube closes by Hamburger-Hamilton (HH) stage 9 (around 24 hours), and the heart tube forms by HH stage 10. Limb buds emerge by HH stage 18 (approximately 50 hours), and organ primordia such as the liver, lungs, and gut become distinct by HH stage 22. Unlike frogs, chick embryos develop within eggs, allowing for a more gradual and coordinated development of organs. The formation of feathers and specialized respiratory systems further extends the organogenesis phase, typically lasting up to 21 days before hatching.
Mouse embryos, as mammals, have the longest organogenesis period among the three species, spanning approximately 10.5 to 18.5 days post-coitum (dpc). The neural tube closes by 8.5 dpc, and the heart begins to form by 8.0 dpc, with a functional heartbeat by 8.5 dpc. Limb buds appear by 9.5 dpc, and major organs such as the liver, lungs, and kidneys undergo rapid differentiation between 10.5 and 14.5 dpc. The extended timeline in mice is attributed to the development of complex mammalian-specific structures, such as the placenta and highly specialized brain regions. This prolonged organogenesis ensures the embryo’s readiness for postnatal life.
Comparatively, frog organogenesis is the fastest, driven by the need for rapid aquatic adaptation, while chick and mouse embryos exhibit more protracted timelines due to their amniotic and mammalian complexities, respectively. Despite these differences, key events such as neural tube closure, heart formation, and limb bud emergence occur in a conserved sequence across species, highlighting shared evolutionary origins. These timelines underscore the balance between developmental speed and complexity, offering a framework for studying vertebrate embryology.
Chicken Egg Count on a Goose Egg
You may want to see also
![]()
Yolk Utilization: Role of yolk in nutrient supply and embryonic growth across species
The role of yolk in embryonic development is a critical aspect of understanding the diverse strategies employed by different species to ensure the growth and survival of their offspring. In the context of frog, chick, and mouse embryology, yolk utilization varies significantly, reflecting the unique reproductive adaptations of each species. Yolk, a nutrient-rich cellular material, serves as a vital energy source and provides essential compounds for the developing embryo, but its contribution differs across these vertebrates.
In frogs, embryogenesis relies heavily on the yolk's nutritional reserves. Frog eggs are typically large and contain a substantial amount of yolk, which is crucial for the early stages of development. The yolk provides lipids, proteins, and other nutrients necessary for cell division and differentiation. As the embryo develops, it utilizes the yolk's resources, and the yolk sac becomes a prominent feature, offering a visual indicator of the embryo's growth. This yolk-dependent strategy allows frogs to produce fewer, larger eggs, ensuring a higher chance of survival for the offspring in aquatic environments.
Chick embryology presents a different scenario, where the yolk's role is equally important but more specialized. Chicken eggs consist of a significant yolk portion, which is not directly consumed by the developing embryo. Instead, the yolk is connected to the embryo via the vitelline membrane and the yolk sac. During incubation, the embryo absorbs nutrients from the yolk, primarily lipids and proteins, through this connection. The yolk's nutrients are crucial for the rapid growth of the chick, especially for the development of organs and tissues. Interestingly, the yolk also plays a role in waste management, as it helps in the temporary storage of embryonic waste products.
Mouse embryology, on the other hand, exhibits a distinct approach to yolk utilization. Mice, being mammals, have a placental connection, which significantly reduces the reliance on yolk for nutrient supply. The yolk sac in mice is primarily involved in the early stages of embryonic development, providing essential nutrients and contributing to the formation of the embryo's blood cells. However, as the placenta develops, it takes over the role of nutrient supply, and the yolk's significance diminishes. This transition highlights the evolutionary adaptation of mammals towards a more direct maternal-fetal nutrient exchange system.
The comparison of these three species reveals a fascinating diversity in yolk utilization strategies. While frogs and chicks depend on yolk for a substantial part of their embryonic nutrition, mice utilize it only temporarily. These variations are closely tied to the reproductive strategies and environmental constraints of each species. Understanding these differences provides valuable insights into the evolutionary pathways that shape the development and survival of vertebrates.
In summary, the study of yolk utilization across species offers a unique perspective on the intricate relationship between reproductive biology and embryonic development. Each species has evolved distinct mechanisms to ensure the successful growth of their offspring, with yolk playing a central or supporting role depending on the specific requirements of the embryo's environment and developmental needs. This knowledge contributes to our broader understanding of the remarkable diversity in the animal kingdom's reproductive strategies.
KFC Bucket Costs: How Much for That Finger Lickin' Goodness?
You may want to see also
Frequently asked questions
Frog embryology development includes fertilization, cleavage, blastula formation, gastrulation, neurulation, and organogenesis. Key stages involve the formation of the blastopore, which becomes the mouth, and the development of the tail and limbs during the tadpole stage.
Chick embryology involves amniotic development, with extraembryonic membranes like the amnion and yolk sac. Unlike frogs, chicks undergo internal fertilization and have a longer gestation period. Chick embryos also develop a more complex body plan with distinct head, trunk, and limb structures early on.
Mouse embryology includes fertilization, implantation, gastrulation, neurulation, and organogenesis. Major milestones are the formation of the primitive streak, neural tube closure, and the development of the three germ layers (ectoderm, mesoderm, endoderm) that give rise to all tissues and organs.
Environmental factors like temperature, oxygen levels, and chemical exposure can significantly impact embryology development. For example, temperature affects sex determination in frogs, while nutrient availability influences chick growth. In mice, maternal diet and stress can alter fetal development and long-term health outcomes.



















