
The development of the chick embryo is a fascinating subject in embryology, offering valuable insights into vertebrate development. One key aspect of this process is the formation of somites, which are paired blocks of mesoderm that give rise to various tissues, including the vertebrae, ribs, and skeletal muscle. Researchers often study the chick embryo to understand the precise timing and mechanisms of somite formation. A common question in this field is how many somites are typically present in a 24-hour chick embryo. This inquiry is crucial as it helps establish developmental milestones and provides a baseline for comparing normal and abnormal embryonic growth. Understanding the number of somites at this stage not only aids in developmental biology research but also has implications for regenerative medicine and congenital disorder studies.
Explore related products






What You'll Learn
![]()
Somite Formation Rate in Chick Embryos
Somite formation in chick embryos is a critical process during early development, marking the segmentation of the mesoderm into repetitive blocks that give rise to vertebral and rib structures. The rate at which somites form is a key parameter in understanding embryonic development, as it reflects the precision and timing of molecular and cellular mechanisms. In chick embryos, somite formation begins around Hamburger-Hamilton (HH) stage 7 to 8, and the process continues at a relatively consistent rate during the first 24 hours of incubation. Research indicates that chick embryos typically form approximately 40 to 50 somites within this 24-hour period, depending on factors such as temperature and genetic background.
The somite formation rate in chick embryos is regulated by the segmentation clock, a molecular oscillator that drives the periodic formation of somites from the presomitic mesoderm (PSM). This clock operates through oscillating gene expression, primarily involving genes like *Lunatic Fringe* (*Lfng*), *Hairy1* (*H1*), and *Delta-like 1* (*Dll1*). The periodicity of the segmentation clock in chick embryos is approximately 90 minutes, meaning a new somite is formed roughly every 90 minutes. This rate is temperature-dependent; for example, at 38°C, the optimal incubation temperature for chick embryos, the segmentation clock maintains its periodicity, ensuring consistent somite formation.
Experimental observations have shown that the number of somites in a 24-hour chick embryo can vary slightly due to biological variability and environmental conditions. However, the average count falls within the range of 40 to 50 somites, with the first somite appearing around 18 to 20 hours post-incubation. This rate is crucial for developmental biologists studying the mechanisms of somitogenesis, as deviations from the normal rate can indicate abnormalities in the segmentation clock or other developmental pathways. For instance, perturbations in the Notch or Wnt signaling pathways can disrupt somite formation, leading to fewer or malformed somites.
To study somite formation rate, researchers often use time-lapse microscopy or molecular markers to track the emergence of somites in real time. Techniques such as in situ hybridization or live imaging with fluorescent reporters allow for precise monitoring of gene expression oscillations and somite boundaries. These methods have confirmed that the rate of somite formation in chick embryos is highly conserved, reflecting the robustness of the underlying molecular mechanisms. Understanding this rate is not only fundamental to developmental biology but also has implications for regenerative medicine and the study of congenital disorders related to vertebral malformations.
In conclusion, the somite formation rate in chick embryos is a tightly regulated process, with approximately 40 to 50 somites formed within the first 24 hours of development. This rate is governed by the segmentation clock, which operates with a periodicity of about 90 minutes. The consistency of somite formation highlights the precision of embryonic development and provides a valuable model for studying the molecular and cellular basis of segmentation. By investigating this process, scientists gain insights into the fundamental principles of developmental biology and the mechanisms that ensure proper tissue patterning in vertebrates.
McDonald's Secret Menu: Chicken Big Mac
You may want to see also
Explore related products






![]()
Molecular Mechanisms of Somite Segmentation
The process of somite segmentation is a fascinating aspect of embryonic development, particularly in the context of the 24-hour chick embryo, which typically exhibits around 8-12 somites during this stage. Somites are the precursor structures that give rise to vertebrae, ribs, and skeletal muscle, making their formation a critical event in vertebrate embryogenesis. The molecular mechanisms underlying somite segmentation involve a complex interplay of genetic oscillations, signaling pathways, and cellular interactions. Central to this process is the segmentation clock, a molecular oscillator that operates in the presomitic mesoderm (PSM) and coordinates the periodic formation of somites. This clock is driven by the cyclic expression of genes such as *Hes7* and *Lunatic Fringe* (*Lfng*), which are part of the Notch signaling pathway. The oscillatory dynamics of these genes create a traveling wave of gene expression that sweeps across the PSM, defining the boundaries of future somites.
The Notch signaling pathway plays a pivotal role in somite segmentation by regulating the synchronization and coordination of oscillations in the PSM cells. Notch signaling is activated through interactions between Delta ligands on neighboring cells and Notch receptors, leading to the cleavage of the Notch intracellular domain (NICD). NICD then translocates to the nucleus, where it activates the transcription of target genes, including *Hes7*. The cyclic activation and repression of *Hes7* create a negative feedback loop that sustains oscillations. Additionally, *Lfng*, a glycosyltransferase, modulates Notch signaling by altering the affinity of Notch receptors for their ligands, further refining the oscillatory pattern. This precise regulation ensures that somites are formed at regular intervals, a process known as somitogenesis.
Another critical pathway involved in somite segmentation is the Wnt signaling pathway, which interacts with Notch signaling to establish the anterior-posterior axis and define somite boundaries. Wnt signaling gradients, particularly involving Wnt3a, create a spatial pattern that influences the oscillations driven by Notch. The interplay between Wnt and Notch pathways is essential for maintaining the periodicity and synchronization of the segmentation clock. Disruptions in either pathway can lead to defects in somite formation, highlighting their interdependence in this process.
Furthermore, the fibroblast growth factor (FGF) signaling pathway contributes to somite segmentation by regulating the progression of the segmentation clock. FGF signaling controls the state of PSM cells, determining whether they remain in an oscillatory state or differentiate into somites. Gradients of FGF activity along the PSM influence the timing and position of somite formation. The integration of FGF signaling with Notch and Wnt pathways ensures that somites are segmented with precise spatial and temporal control.
Lastly, the role of retinoic acid (RA) signaling in somite segmentation cannot be overlooked. RA, a derivative of vitamin A, acts as a morphogen that patterns the anterior-posterior axis and influences the segmentation clock. RA signaling modulates the expression of segmentation clock genes, such as *Hes7*, and coordinates the transition of PSM cells into somites. The graded distribution of RA along the embryonic axis provides positional information that is crucial for the accurate segmentation of somites.
In summary, the molecular mechanisms of somite segmentation in the 24-hour chick embryo involve a highly coordinated network of signaling pathways, including Notch, Wnt, FGF, and retinoic acid. These pathways work in concert to establish and maintain the segmentation clock, ensuring the periodic and precise formation of somites. Understanding these mechanisms not only sheds light on the developmental processes of vertebrates but also provides insights into the broader principles of pattern formation in biology.
Dave's Hot Chicken Hiring Age: What You Need to Know
You may want to see also
Explore related products






![]()
Role of Oscillatory Genes in Somitogenesis
The process of somitogenesis, where somites (the precursors to vertebrae and associated muscles) form in a chick embryo, is a highly regulated and dynamic event. In a 24-hour chick embryo, approximately 8-12 somites are typically present, depending on the specific developmental stage and environmental conditions. This precise segmentation is governed by a complex interplay of genetic and molecular mechanisms, with oscillatory genes playing a pivotal role. These genes, characterized by their cyclic expression patterns, create a molecular clock that drives the rhythmic formation of somites from the presomitic mesoderm (PSM).
Oscillatory genes, such as those in the Notch, Wnt, and FGF signaling pathways, establish a segmentation clock that coordinates the periodic release of cells from the PSM into newly formed somites. The Notch pathway, for instance, is crucial for maintaining the oscillatory dynamics in the PSM. Genes like *Lunatic Fringe* (*Lfng*) and *Hes7* (in mammals, homologous to *c-Hey2* in chicks) exhibit cyclic expression, creating waves of activity that travel through the PSM. These oscillations ensure that cells in the PSM are synchronized, allowing them to differentiate into somites at regular intervals. Disruption of these oscillatory patterns can lead to defects in somite formation, highlighting their essential role in somitogenesis.
The Wnt and FGF pathways also contribute to the oscillatory mechanism by regulating the activity of the Notch pathway and maintaining the proliferative and undifferentiated state of PSM cells. Wnt signaling, in particular, is involved in the posteriorization of the embryo, ensuring that the PSM continues to produce new somites. FGF signaling, on the other hand, modulates the speed of the segmentation clock, influencing the periodicity of somite formation. Together, these pathways create a robust oscillatory system that is both precise and adaptable, ensuring the correct number of somites form within the 24-hour window in chick development.
Mathematical modeling has further elucidated the role of oscillatory genes in somitogenesis, demonstrating how their cyclic expression patterns translate into spatial and temporal patterns of somite formation. These models suggest that the oscillations in gene expression create a traveling wave of signals that sweep across the PSM, with each wave corresponding to the formation of a new somite. The frequency and synchronization of these waves are critical for determining the number and size of somites, which in a 24-hour chick embryo, typically results in 8-12 somites. This precision is a testament to the finely tuned nature of the oscillatory gene network.
In conclusion, oscillatory genes are central to the process of somitogenesis, driving the rhythmic and coordinated formation of somites in the developing chick embryo. Through their cyclic expression and interaction with key signaling pathways, these genes establish a segmentation clock that ensures the correct number of somites form within the first 24 hours of development. Understanding the role of these genes not only sheds light on the molecular basis of embryonic patterning but also provides insights into potential mechanisms underlying developmental disorders related to somite formation. The study of oscillatory genes in somitogenesis thus remains a critical area of research in developmental biology.
Tasty Chicken Meat: Which Part is Best?
You may want to see also
Explore related products






![]()
Impact of Temperature on Somite Development
Temperature plays a critical role in the development of somites, the segmental structures that give rise to vertebrae, ribs, and associated muscles in vertebrates, including chicks. Research has shown that temperature can significantly influence the rate and pattern of somite formation during embryonic development. In the context of a 24-hour chick embryo, the number of somites formed is highly sensitive to temperature variations. Typically, under standard incubation conditions (around 37.5°C), a chick embryo develops approximately 8-12 somites within the first 24 hours. However, deviations from this optimal temperature can lead to alterations in somite number, morphology, and segmentation clock timing.
Elevated temperatures, for instance, can accelerate the pace of somite formation by increasing the speed of molecular oscillations in the segmentation clock, a genetic oscillator that controls the periodic formation of somites. This results in a higher number of somites within the same time frame. For example, studies have demonstrated that chick embryos incubated at 39°C may exhibit up to 14-16 somites within 24 hours due to the hastened segmentation process. Conversely, lower temperatures slow down the segmentation clock, leading to fewer somites. At 35°C, a chick embryo might develop only 6-8 somites in 24 hours, as the reduced metabolic rate delays the formation of each somite.
The impact of temperature on somite development is not limited to the number of somites but also extends to their size and organization. High temperatures can cause somites to be smaller and less distinct, potentially disrupting the proper differentiation of tissues derived from somites. Conversely, lower temperatures may result in larger but fewer somites, which could lead to abnormalities in axial skeleton development. These temperature-induced changes are mediated by temperature-sensitive proteins and enzymes involved in the Notch and Wnt signaling pathways, which are crucial for somite segmentation.
Practical implications of temperature effects on somite development are particularly relevant in poultry farming and embryological research. In industrial hatcheries, maintaining precise incubation temperatures is essential to ensure uniform and healthy chick development. Even slight temperature fluctuations during critical developmental windows can lead to batch inconsistencies, affecting the quality and viability of chicks. For researchers, understanding temperature-somite interactions provides insights into the mechanisms of embryonic patterning and the evolutionary conservation of temperature sensitivity in developmental processes.
In conclusion, temperature acts as a key regulator of somite development in chick embryos, influencing both the quantity and quality of somites formed within the first 24 hours. Optimal temperature ensures the proper progression of the segmentation clock and the formation of a normal number of somites, while deviations can lead to developmental abnormalities. This sensitivity to temperature highlights the intricate balance required for embryonic development and underscores the importance of controlled environmental conditions in both research and agricultural settings.
Raw Chicken Emergency: Quick Steps to Take After Accidental Consumption
You may want to see also
Explore related products





![]()
Comparison of Somite Counts Across Species
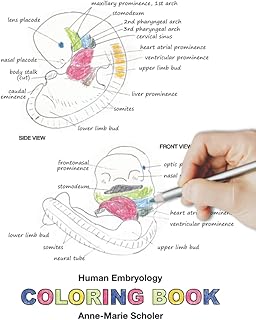
The number of somites present in a developing embryo is a critical indicator of its stage of development, and this count varies significantly across species. In the context of a 24-hour chick embryo, research indicates that approximately 8-12 somites are typically present, depending on the exact timing and environmental conditions. This range serves as a baseline for comparing somite counts in other species, highlighting both similarities and divergences in early developmental processes. Understanding these variations is essential for studying evolutionary biology, developmental mechanisms, and interspecies differences in embryogenesis.
When comparing the 24-hour chick embryo to other avian species, such as quail or duck, the somite counts are relatively consistent, often falling within a similar range of 8-12 somites. This similarity suggests conserved developmental pathways among closely related species. However, when extending the comparison to non-avian species, the differences become more pronounced. For instance, a 24-hour mouse embryo typically exhibits a higher somite count, ranging from 18 to 22 somites. This disparity reflects the faster pace of mouse embryogenesis compared to chicks, despite both being amniotes. Such comparisons underscore the influence of evolutionary adaptations on developmental timing and morphology.
In contrast, examining somite counts in non-amniote species, such as zebrafish, reveals even more striking differences. A 24-hour zebrafish embryo has already formed all 30-32 somites, a process that occurs within the first 12-16 hours of development. This rapid somite formation is attributed to the zebrafish's shorter gestation period and distinct developmental mechanisms. These cross-species comparisons highlight the diversity in somite formation rates and the underlying genetic and environmental factors that drive these variations.
Further analysis of somite counts across species also provides insights into the modularity of vertebrate development. For example, while the total number of somites differs, the process of somitogenesis—the formation of somites from the presomitic mesoderm—shares conserved molecular pathways across vertebrates. This includes the oscillatory expression of genes in the segmentation clock, which regulates the periodic formation of somites. Despite these commonalities, the timing and duration of somitogenesis vary, leading to species-specific somite counts at equivalent developmental stages.
In conclusion, comparing somite counts across species, such as the 24-hour chick embryo, reveals both conserved and divergent aspects of early development. While closely related species like quail and duck exhibit similar somite counts, more distantly related species like mice and zebrafish show accelerated somite formation. These variations reflect evolutionary adaptations in developmental timing and morphology, while also highlighting the modular nature of somitogenesis. Such comparisons are invaluable for understanding the evolutionary plasticity of developmental processes and the mechanisms that shape vertebrate embryogenesis.
Uncovering the Source: Where Does Church's Chicken Truly Originate?
You may want to see also
Frequently asked questions
A 24-hour chick embryo usually has around 12-14 somites, depending on the exact developmental stage and environmental conditions.
The number of somites in a 24-hour chick embryo is a critical indicator of developmental progress, as somites are the precursors to vertebrae, ribs, and skeletal muscle, and their count reflects the embryo's growth and health.
Yes, the number of somites can vary slightly due to factors like temperature, genetic background, and experimental conditions, but 12-14 somites is the typical range for this stage.































