The closure of the chick neural tube during embryonic development is a critical event that marks the formation of the central nervous system. In chick embryos, this process typically occurs between stages 8 and 12 of Hamburger and Hamilton (HH) development, which corresponds to approximately 24 to 30 hours after fertilization. Neural tube closure begins at the future midbrain-hindbrain boundary and progresses both cranially and caudally, eventually sealing to form the brain and spinal cord. This tightly regulated process involves coordinated cellular movements, including convergent extension and zippering, and is influenced by genetic and molecular signals such as the planar cell polarity pathway and Wnt signaling. Proper closure is essential, as disruptions can lead to neural tube defects, highlighting its significance in developmental biology and teratology.
| Characteristics | Values |
|---|---|
| Species | Chick (Gallus gallus domesticus) |
| Neural Tube Closure Stage | Hamburger-Hamilton (HH) stages 5-9 |
| Time Frame | Approximately 24-36 hours post-fertilization (depending on incubation conditions) |
| Closure Initiation | Begins at the future midbrain-hindbrain boundary (isthmus) |
| Closure Progression | Zipper-like closure, moving both cranially (head) and caudally (tail) |
| Completion | Fully closed by HH stage 9 |
| Key Molecular Signals | BMP, Wnt, Shh, and planar cell polarity (PCP) pathway components |
| Morphological Changes | Neural folds elevate, fuse, and form a closed neural tube |
| Temperature Dependence | Optimal incubation at 37.5°C; deviations can delay or disrupt closure |
| Clinical Relevance | Defects in closure lead to neural tube defects (NTDs) in avian models |
| Research Significance | Widely studied as a model for vertebrate neural tube closure mechanisms |
Explore related products






What You'll Learn
- Timing in Chick Embryos: Neural tube closure occurs around stages 9-10, approximately 24-30 hours post-fertilization
- Closure Mechanism: Zipper-like process starts at future midbrain, progresses rostrally and caudally simultaneously
- Key Molecular Signals: Involves planar cell polarity (PCP) pathway, Wnt, and Shh signaling for coordinated closure
- Morphological Changes: Convergent extension and cell shape changes drive tissue movement during closure
- Defects and Causes: Failure results in neural tube defects, linked to genetic or environmental factors
![]()
Timing in Chick Embryos: Neural tube closure occurs around stages 9-10, approximately 24-30 hours post-fertilization
The timing of neural tube closure in chick embryos is a critical event in early development, marking a significant milestone in the formation of the central nervous system. This process occurs during a precise window, specifically around stages 9-10 of embryonic development, which corresponds to approximately 24-30 hours post-fertilization. At this stage, the neural folds, which have previously elevated along the midline of the embryo, begin to converge and fuse, transforming the open neural plate into a closed neural tube. This closure is essential for the subsequent differentiation and organization of neural tissues.
During stages 9-10, the chick embryo undergoes rapid morphological changes, and the timing of neural tube closure is tightly regulated by genetic and molecular signals. The process begins at the future hindbrain region and progresses both cranially (toward the head) and caudally (toward the tail) in a zipper-like fashion. This coordinated movement ensures that the neural tube closes completely, preventing defects such as spina bifida or anencephaly, which can arise from incomplete closure. The precise timing of this event highlights the importance of developmental staging in chick embryology, as even slight deviations can impact the embryo's viability.
The 24- to 30-hour post-fertilization window is a period of heightened cellular activity, with epithelial-to-mesenchymal transitions and cytoskeletal rearrangements driving the fusion of the neural folds. Researchers often use this timeframe as a reference point for studying the molecular mechanisms underlying neurulation. For example, experiments may focus on the role of proteins like sonic hedgehog (Shh) or bone morphogenetic proteins (BMPs), which are known to regulate neural tube formation. Understanding the timing of closure in chick embryos also provides insights into comparative developmental biology, as similar processes occur in other vertebrates, including humans.
From a practical standpoint, the timing of neural tube closure in chick embryos is crucial for experimental design in developmental biology. Researchers must carefully synchronize embryonic stages to study specific aspects of neurulation, often using techniques like in ovo electroporation or chemical perturbations during this critical window. The chick model is particularly valuable due to its accessibility and the ability to manipulate the embryo at early stages. By focusing on the 24- to 30-hour period, scientists can gain a deeper understanding of the cellular and molecular events that ensure proper neural tube closure.
In summary, neural tube closure in chick embryos occurs around stages 9-10, approximately 24-30 hours post-fertilization, representing a pivotal moment in early development. This tightly regulated process involves the fusion of neural folds and is essential for the formation of the central nervous system. The precise timing of this event not only underscores its biological significance but also provides a critical framework for research in developmental biology. By studying this process in chick embryos, scientists can uncover fundamental principles of neurulation that have broader implications for understanding and addressing neural tube defects in various species.
Discovering Panama's Twist on the Classic Chicken Piccata Recipe
You may want to see also
Explore related products






![]()
Closure Mechanism: Zipper-like process starts at future midbrain, progresses rostrally and caudally simultaneously
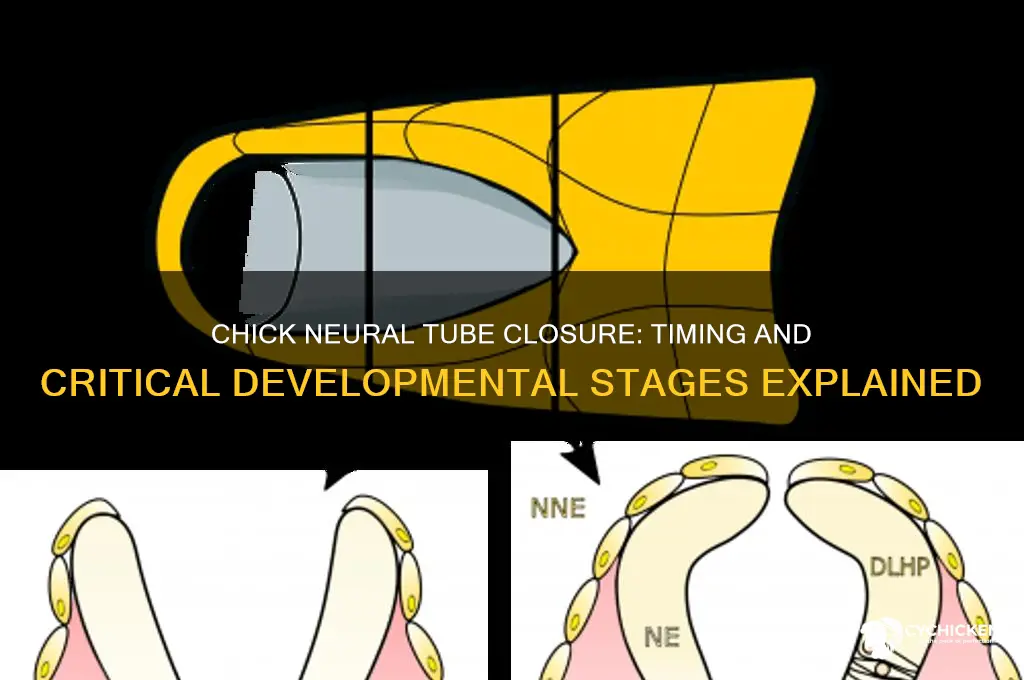
The closure of the chick neural tube during embryonic development is a highly coordinated and dynamic process, essential for the proper formation of the central nervous system. One of the most fascinating aspects of this process is the closure mechanism, which resembles a zipper-like action. This mechanism initiates at the future midbrain region, a critical site that marks the beginning of neural tube closure. From this point, the process progresses simultaneously in two directions: rostrally (toward the head) and caudally (toward the tail). This bidirectional movement ensures that the neural folds converge and fuse efficiently, sealing the neural tube along its entire length.
The zipper-like closure is driven by intricate cellular and molecular events. At the future midbrain, the neural folds elevate and approach each other due to changes in cell shape, adhesion, and extracellular matrix remodeling. This elevation is facilitated by the coordinated activity of actin and microtubule cytoskeletons within the neuroepithelial cells. As the neural folds rise, they begin to fuse at the midline, forming a seamless epithelial structure. This fusion is mediated by tight junctions and adherens junctions, which strengthen the connection between cells and prevent the ingress of external fluids or tissues.
As closure progresses rostrally and caudally, the process is tightly regulated by signaling pathways such as Wnt, BMP, and Shh, which ensure proper cell differentiation and tissue morphogenesis. The rostral progression is particularly critical, as it involves the closure of the anterior neuropore, which gives rise to the forebrain and midbrain structures. Simultaneously, the caudal progression seals the posterior neuropore, forming the spinal cord. The simultaneous nature of this process is crucial for preventing neural tube defects, as delays or disruptions in either direction can lead to severe developmental abnormalities.
Mechanistically, the zipper-like closure relies on the coordinated movement of cells within the neural plate. Cells at the leading edge of the neural folds undergo apical constriction, reducing their height and pulling the folds closer together. This constriction is driven by acto-myosin contractility, where actin filaments and myosin motors generate the necessary force for tissue bending and fusion. Additionally, planar cell polarity (PCP) pathways play a key role in directing cell movements and ensuring that the neural folds converge uniformly along the rostro-caudal axis.
In the chick embryo, this closure process typically occurs around Hamburger-Hamilton stage 8 to 10, which corresponds to approximately 24 to 30 hours after fertilization. The precise timing and coordination of this mechanism are vital, as any deviation can result in neural tube defects such as anencephaly or spina bifida. Understanding the zipper-like closure process in chicks provides valuable insights into the conserved mechanisms of neural tube formation across vertebrates, including humans, and highlights the importance of developmental biology in studying congenital disorders.
Mastering Chicken Fabrication: A Detailed Step-by-Step Guide for Beginners
You may want to see also
Explore related products






![]()
Key Molecular Signals: Involves planar cell polarity (PCP) pathway, Wnt, and Shh signaling for coordinated closure
The closure of the chick neural tube during embryonic development is a highly coordinated process that occurs around Hamburger-Hamilton stage 8-10, approximately 24-30 hours after incubation. This critical event relies on precise molecular signaling pathways to ensure proper morphogenesis. Among these, the planar cell polarity (PCP) pathway, Wnt signaling, and Sonic Hedgehog (Shh) signaling play pivotal roles in orchestrating the cellular movements and tissue reshaping required for neural tube closure. These pathways interact dynamically to regulate cell adhesion, cytoskeletal reorganization, and tissue convergence, ensuring the seamless fusion of the neural folds.
The PCP pathway is essential for establishing tissue-wide polarity and coordinating directed cell movements during neural tube closure. In chicks, PCP signaling ensures that cells align and move in a synchronized manner along the plane of the neural plate. Core PCP proteins, such as Vangl2 and Fz6, localize asymmetrically at the cell membrane, guiding the actin cytoskeleton and cell protrusions. This polarization enables cells to migrate collectively toward the dorsal midline, facilitating the elevation and fusion of the neural folds. Disruption of PCP components in chick embryos results in open neural tube defects, underscoring its critical role in this process.
Wnt signaling acts in concert with the PCP pathway to regulate neural tube closure in chick embryos. Wnt ligands, particularly Wnt5a, activate the non-canonical Wnt/PCP pathway, promoting cytoskeletal rearrangements and cell migration. Additionally, Wnt signaling modulates cell proliferation and differentiation in the neural plate border, ensuring a sufficient pool of cells for closure. Studies in chick models have shown that Wnt5a gradients guide convergent extension movements, a key process during neural tube closure. Misregulation of Wnt signaling leads to defects in neural fold fusion, highlighting its importance in both PCP-dependent and -independent mechanisms.
Sonic Hedgehog (Shh) signaling is another critical regulator of neural tube closure, primarily through its role in patterning the neural tube and maintaining tissue integrity. Shh, secreted from the notochord and floor plate, establishes a ventral-to-dorsal gradient that influences cell fate and morphogenesis. In chick embryos, Shh signaling ensures proper differentiation of the neural plate border cells, which are essential for initiating closure. Furthermore, Shh coordinates with Wnt and PCP pathways to regulate the expression of adhesion molecules like N-cadherin, which are vital for cell-cell interactions during tissue convergence. Perturbation of Shh signaling in chick models disrupts neural tube closure, emphasizing its interplay with other pathways.
In summary, the closure of the chick neural tube is a tightly regulated process that depends on the integration of PCP, Wnt, and Shh signaling. These pathways work in tandem to control cell polarity, migration, and adhesion, ensuring the precise convergence and elevation of neural folds. Understanding their molecular interplay provides critical insights into the developmental mechanisms underlying neural tube closure and offers potential targets for preventing neural tube defects in humans.
Spanish Farmer's Secret to Happy Chickens
You may want to see also
Explore related products






![]()
Morphological Changes: Convergent extension and cell shape changes drive tissue movement during closure
In the context of chick neural tube closure, which occurs around embryonic day 2 (E2) to E3, morphological changes play a pivotal role in driving tissue movement. Among these, convergent extension and cell shape changes are the primary mechanisms that facilitate the dynamic rearrangement of cells during this critical developmental process. Convergent extension involves the coordinated movement of cells toward the midline (convergence) and their subsequent elongation along the anterior-posterior axis (extension). This process is essential for narrowing and elongating the neural plate, setting the stage for neural tube closure. In the chick embryo, convergent extension begins shortly after gastrulation and continues as the neural folds elevate and fuse.
During convergent extension, cells undergo significant shape changes that enable their directed movement. Initially, cells in the neural plate are polygonal and tightly packed. As closure progresses, these cells become elongated and polarized, aligning themselves along the axis of extension. This polarization is driven by the reorganization of the cytoskeleton, particularly actin and microtubules, which generate the forces necessary for cell intercalation and migration. The planar cell polarity (PCP) pathway is a key regulator of this process, ensuring that cells respond appropriately to directional cues and maintain coordinated movement.
Cell intercalation, a critical component of convergent extension, involves the rearrangement of cells relative to their neighbors. In the chick neural plate, medial-lateral intercalation brings cells closer to the midline, while anterior-posterior intercalation contributes to tissue elongation. These movements are facilitated by the formation and contraction of adherens junctions, which allow cells to slide past one another while maintaining tissue integrity. The precise regulation of these junctions, along with the activity of motor proteins like non-muscle myosin II, ensures that tissue deformation occurs in a controlled and directed manner.
The elevation of the neural folds, a subsequent step in neural tube closure, is also driven by morphological changes in cell shape and arrangement. As the neural plate narrows due to convergent extension, the edges of the plate, or neural folds, begin to bend upward. This bending is accompanied by apical constriction, where cells at the apical surface of the neuroepithelium reduce their height, causing the tissue to curve inward. Apical constriction is mediated by actomyosin networks, which generate contractile forces that pull the apical surfaces of cells together, contributing to fold elevation.
Finally, the fusion of the neural folds at the dorsal midline completes the process of neural tube closure. This step requires the precise alignment and adhesion of cells from opposing folds, a process that is again dependent on cell shape changes and cytoskeletal dynamics. The coordinated activity of convergent extension, cell intercalation, and apical constriction ensures that the neural tube closes efficiently and accurately, preventing developmental defects such as spina bifida. Understanding these morphological changes provides critical insights into the mechanisms driving tissue morphogenesis during chick neural tube closure.
Crockpot Cooking: Is Raw Chicken Safe?
You may want to see also
Explore related products



![]()
Defects and Causes: Failure results in neural tube defects, linked to genetic or environmental factors
In the context of chick embryonic development, the neural tube closure typically occurs around embryonic day 2.5 to 4.5, depending on the specific region of the tube. This critical process involves the fusion of neural folds to form a hollow tube, which subsequently gives rise to the central nervous system. Failure of the neural tube to close properly during this narrow developmental window results in neural tube defects (NTDs), a group of severe congenital malformations. These defects can manifest as anencephaly, where the brain and skull are incompletely formed, or spina bifida, characterized by incomplete closure of the spinal cord and vertebrae. Understanding the timing and mechanisms of neural tube closure is essential for identifying the factors that contribute to these developmental abnormalities.
Genetic Factors: NTDs have a strong genetic component, with both inherited and de novo mutations playing significant roles. For instance, mutations in genes involved in the planar cell polarity (PCP) pathway, such as *VANGL1* and *CELSR1*, disrupt the coordinated cell movements necessary for neural tube closure. Similarly, defects in folate metabolism genes, like *MTHFR*, impair the availability of methyl groups required for DNA synthesis and cellular proliferation during neurulation. Familial studies have shown that a history of NTDs in first-degree relatives increases the risk of recurrence, highlighting the heritable nature of these defects. Genetic screening and counseling are crucial for families with a history of NTDs to assess and mitigate risks in subsequent pregnancies.
Environmental Factors: Environmental factors, particularly maternal nutrition and exposure to teratogens, are closely linked to NTDs. Folate deficiency during early pregnancy is a well-established risk factor, as folate is critical for nucleotide synthesis and DNA methylation, processes essential for neural tube closure. Maternal diabetes, obesity, and hyperthermia (e.g., from fever or sauna use) during early gestation also increase the risk of NTDs by disrupting embryonic development. Exposure to certain medications, such as valproic acid and carbamazepine, and environmental toxins like mycotoxins and pesticides, can interfere with cellular signaling pathways involved in neurulation. Public health interventions, including folic acid supplementation and prenatal care, have significantly reduced the incidence of NTDs in populations with access to these resources.
Gene-Environment Interactions: The etiology of NTDs often involves complex interactions between genetic predisposition and environmental triggers. For example, individuals with *MTHFR* mutations may be more susceptible to NTDs when maternal folate intake is insufficient, as the mutation impairs the body's ability to utilize folate effectively. Similarly, maternal diabetes exacerbates the risk of NTDs in genetically susceptible embryos by altering glucose metabolism and oxidative stress levels. Understanding these interactions is critical for developing targeted prevention strategies that address both genetic vulnerabilities and modifiable environmental risks.
Prevention and Research Directions: Preventive measures for NTDs focus on addressing known risk factors, such as promoting periconceptional folic acid supplementation and managing maternal health conditions like diabetes. Advances in genomic research and animal models, including chick embryos, have enhanced our understanding of the molecular mechanisms underlying neural tube closure and its defects. Future research should aim to identify additional genetic markers, explore the role of epigenetic factors, and develop personalized interventions to reduce the global burden of NTDs. Public awareness campaigns and improved access to prenatal care remain essential for translating scientific knowledge into effective prevention strategies.
Would You Rather: Chicken Fights or Avoidance Strategies?
You may want to see also
Frequently asked questions
The chick neural tube typically closes between stages 9 and 10 of Hamburger-Hamilton (HH) development, which corresponds to approximately 24 to 30 hours after fertilization.
Neural tube closure in chick embryos is influenced by genetic factors, signaling pathways (such as Wnt, BMP, and Shh), and mechanical forces. Proper closure also depends on the coordinated movement of cells and tissues during gastrulation.
Failure of the neural tube to close properly in chick embryos can lead to neural tube defects (NTDs), which may result in severe developmental abnormalities affecting the brain and spinal cord. These defects can be caused by genetic mutations, environmental factors, or disruptions in signaling pathways.



















