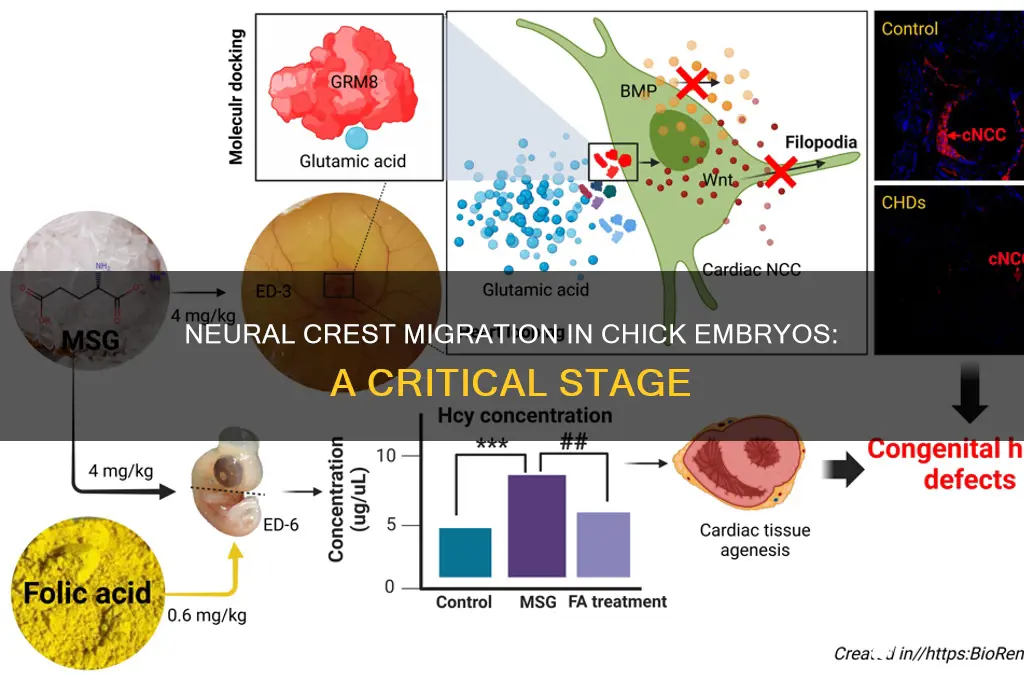
Neural crest cells are a group of cells that delaminate from the dorsal neural tube and migrate extensively throughout the developing embryo. In chicks, neural crest cells migrate to form the cranial and trunk neural crest. The cranial neural crest gives rise to the jaws, teeth, and facial cartilage, while the trunk neural crest forms the dorsal root ganglia and the sympathetic chain. The migration of neural crest cells occurs during embryogenesis and is guided by various signals and molecules, such as semaphorin and integrins. Abnormalities in neural crest development can lead to neurocristopathies, including conditions like DiGeorge syndrome, frontonasal dysplasia, and Waardenburg-Shah syndrome. Understanding the mechanisms of neural crest migration is crucial for advancing our knowledge of human diseases and vertebrate evolution.
| Characteristics | Values |
|---|---|
| Neural crest cells | Originate at the dorsalmost region of the neural tube |
| Migrate extensively to generate different cell types | |
| Undergo a process called delamination | |
| Arise from the dorsal neural tube | |
| Migrate during embryogenesis | |
| Cells remain closely associated with the anterior half of the DM of developing somites as they travel | |
| Neural crest cells express β1 and α4 integrins | |
| Cells that migrate through the rostral half of somites differentiate into sensory and sympathetic neurons of the peripheral nervous system | |
| The other main route neural crest cells take is dorsolaterally between the epidermis and the dermamyotome | |
| Neural crest cells contribute to the peripheral nervous system | |
| Neural crest cells migrate into the pharyngeal arches | |
| Neural crest cells form ectomesenchyme | |
| Neural crest cells contribute to the cardiac neural crest | |
| Neural crest cells contribute to the cranial neural crest | |
| Neural crest cells contribute to the trunk neural crest | |
| Neural crest cells contribute to the sacral neural crest | |
| Neural crest cells contribute to the vagal neural crest |
Explore related products






What You'll Learn
- Neural crest cells migrate from rhombomeres 1 and 2 to form the jawbones
- Neural crest cells migrate into pharyngeal arches.
- Neural crest cells migrate dorsolaterally between the epidermis and dermamyotome
- Neural crest cells undergo delamination, separating from surrounding tissue
- Trunk neural crest cells migrate ventrolaterally through the sclerotome
![]()
Neural crest cells migrate from rhombomeres 1 and 2 to form the jawbones
Neural crest cells are formed along the anteroposterior axis of the embryo, from the posterior diencephalon to the lumbosacral region. They can be grouped into four subpopulations based on their position: cranial, vagal, trunk, or sacral. The cranial neural crest, which includes cells formed between the forebrain and the sixth rhombomere of the hindbrain, is particularly important in the development of the face and jaws.
The hindbrain is segmented along the anterior-posterior axis into compartments called rhombomeres, which play a vital role in the patterning of branchiomotor nerves and cranial ganglia. The neural crest streams are formed adjacent to even-numbered rhombomeres, resulting in a segmented pattern. The first stream, also called the trigeminal neural crest, originates from the midbrain and rhombomeres 1 and 2 (r1-2). These neural crest cells migrate to the first pharyngeal (mandibular) arch, forming the jawbones, as well as the incus and malleus bones of the middle ear.
The migration of neural crest cells occurs through a process called delamination, which involves a full or partial epithelial-mesenchymal transition (EMT). During delamination, neural crest cells separate from the surrounding tissue and undergo a change from an epithelial to a mesenchymal phenotype. In chick embryos, delamination occurs during the fusion of the neural fold.
In chick embryos, semaphorin acts in the cephalic region to guide neural crest cells through the pharyngeal arches. Neural crest cells express β1 and α4 integrins, which allow them to bind and interact with collagen, laminin, and fibronectin of the extracellular matrix as they migrate. The migration of neural crest cells is also influenced by repulsive signaling, such as semaphorin-neuropilin and EphB signaling, which guide the cells through the rostral half of somites.
The development of the neural crest is directed by a gene regulatory network, which consists of interacting signals, transcription factors, and downstream effector genes that determine cell characteristics such as multipotency and migratory capabilities. Abnormalities in neural crest development can lead to neurocristopathies, including conditions such as DiGeorge syndrome, Waardenburg-Shah syndrome, and frontonasal dysplasia.
Free-Range Chicken Law: Oregon's New Move
You may want to see also
Explore related products






![]()
Neural crest cells migrate into pharyngeal arches.
Neural crest cells are highly migratory and have the capacity to differentiate into a wide range of cell types. They originate at the dorsalmost region of the neural tube and migrate extensively to form a variety of cell types, including neurons, glial cells, and pigment-containing cells of the epidermis. In chicks, neural crest cells play a crucial role in the development of the pharyngeal arches, which are a series of bulges on the lateral surface of the head that serve both feeding and respiratory functions.
The process of neural crest cell migration in chicks involves specific pathways and interactions with other tissues. The chick cranial neural crest cells migrate from regions anterior to rhombomere 6, taking one of three major pathways. The first pathway involves cells from rhombomeres 1 and 2 migrating to the first pharyngeal (mandibular) arch, where they contribute to the formation of the jawbones, incus, and malleus bones of the ear. The expanding epidermis pulls these cells to form the frontonasal process, which gives rise to the bones of the face.
The second migratory pathway is taken by cells from rhombomere 4, which populate the second pharyngeal arch and form the hyoid cartilage of the neck. Additionally, the cardiac neural crest, located between the cranial and trunk neural crests, contributes to the development of the third, fourth, and sixth pharyngeal arches. These cells migrate into the arches and generate the endothelium of the aortic arch arteries and the septum between the aorta and pulmonary artery.
The migration of neural crest cells into the pharyngeal arches is guided by specific signals and interactions. In chick embryos, semaphorin acts in the cephalic region to guide neural crest cells through the pharyngeal arches. These cells express β1 and α4 integrins, which allow for binding and guided interaction with collagen, laminin, and fibronectin of the extracellular matrix as they migrate. The migration process is also influenced by the rate of proliferation in the dorsal neural tube, and abnormalities in this region can impact the migration of neural crest cells, leading to abnormalities in the pharyngeal arch artery and outflow tract.
The development of the pharyngeal arches is a complex process that involves the coordination of multiple signals and genes. The pharyngeal arches give rise to various structures in the head and neck, and abnormalities in their development can lead to conditions such as Treacher Collins syndrome, which results in mid and lower face abnormalities. Understanding the mechanisms of neural crest migration and their contributions to the pharyngeal arches is crucial for comprehending vertebrate evolution and human pathology.
Famous Queen Bees in Chicken Girls Season 3
You may want to see also
Explore related products






![]()
Neural crest cells migrate dorsolaterally between the epidermis and dermamyotome
Neural crest cells are formed transiently between the epidermal ectoderm and neural plate during vertebrate development. They give rise to a diverse range of cell lineages, including melanocytes, craniofacial cartilage and bone, smooth muscle, dentin, peripheral and enteric neurons, adrenal medulla, and glia. The neural crest is specified at the border of the neural plate and the non-neural ectoderm after gastrulation. During neurulation, the neural folds (borders of the neural plate) converge at the dorsal midline to form the neural tube. Neural crest cells then undergo an epithelial-to-mesenchymal transition, delaminating from the neuroepithelium and migrating peripherally, where they differentiate into various cell types.
In chick embryos, neural crest cells migrate through the pharyngeal arches, forming the ectomesenchyme and contributing to tissues typically derived from mesoderm, such as cartilage, bone, and connective tissue. The migration of neural crest cells in chicks has been studied using techniques like radioisotopic labeling and quail-chick marking systems, allowing researchers to visualize and track their movement during embryonic development.
The migration pathways of neural crest cells differ depending on their location in the embryo. In the trunk region, neural crest cells initially migrate in the intersomitic spaces and then through the anterior half of the somite along the basal lamina of the dermamyotome. After 24 hours, they switch to the dorsolateral path, migrating dorsolaterally between the epidermis and dermamyotome. This migration is influenced by molecular changes in the neural crest cells themselves, such as the upregulation of specific receptors and the expression of proteins like Slug, RhoB, and EphB2.
The dorsolateral migration of neural crest cells is guided by molecular signals. For example, in mice, melanoblasts depend on the Steel factor produced by the dermamyotome for their dispersal onto the dorsolateral path. They express the receptor c-kit before embarking on this path. However, in avian embryos, c-kit is expressed by melanoblasts after they have migrated dorsolaterally, and the Steel factor is produced by the ectoderm rather than the dermamyotome. Ephrin signaling also plays a crucial role in directing neural crest cell migration. Inhibiting ephrin signaling can lead to the inappropriate invasion of neural and glial precursors into the dorsolateral space. Interestingly, ephrins stimulate the migration of melanoblasts, and this behavior is receptor-specific, with EphB2 regulating the migration response.
When to Euthanize an Injured Chicken: Signs and Symptoms
You may want to see also
Explore related products






![]()
Neural crest cells undergo delamination, separating from surrounding tissue
Neural crest cells are multipotent stem cells located at the side of the neural tube, proximal to the epidermal layer. They are formed after neurulation, when the neural folds converge at the dorsal midline to form the neural tube. The neural crest cells originate from the dorsalmost region of the neural tube, at the border between the neural plate and the non-neural ectoderm.
For neural crest cells to begin their migration, they must undergo a process called delamination, which involves a full or partial epithelial-mesenchymal transition (EMT). Delamination is the separation of tissue into different populations, specifically neural crest cells separating from the surrounding tissue. This process is triggered by a BMP/Wnt cascade that induces the expression of EMT-promoting transcription factors such as SNAI2 and FOXD3.
Prior to delamination, neural crest cells are anchored to neighbouring cells by tight junction proteins such as occludin and cell adhesion molecules such as NCAM and N-Cadherin. Dorsally expressed BMPs initiate delamination by inducing the expression of zinc finger protein transcription factors like snail, slug, and twist. These factors directly contribute to the epithelial-mesenchymal transition by reducing the expression of occludin and N-Cadherin, and by promoting the modification of NCAMs with polysialic acid residues to decrease adhesiveness.
The process of delamination is critical for neural crest development, as the nascent crest cells undergo drastic changes in a short period of time. They must alter their cell- and substrate-adhesion properties, lose cell polarity, and activate the locomotory machinery while continuing to proliferate, survive, and maintain a pool of precursors in the neural epithelium. Once released from the neural tube, neural crest cells must adapt to a new and rapidly changing environment, interpreting cues that guide them to appropriate target sites and prevent aberrant distribution.
The study of neural crest cell migration and delamination has been enhanced by cell-labelling techniques, such as the quail-chick marking system devised by Nicole Le Douarin in 1969. This technique, using chimeras generated through transplantation, allowed researchers to distinguish neural crest cells of one species from the surrounding tissue of another.
Quail vs. Chicken: Understanding Their Size Difference
You may want to see also
Explore related products






![]()
Trunk neural crest cells migrate ventrolaterally through the sclerotome
Neural crest cells are formed along most of the anteroposterior axis of the embryo, from the posterior diencephalon to the lumbosacral region of the embryo. They can be grouped into four distinct subpopulations according to the position where they are specified: cranial, vagal, trunk, or sacral. The trunk neural crest gives rise to two populations of cells.
The first group of cells are fated to become melanocytes and migrate dorsolaterally into the ectoderm towards the ventral midline. The second group of cells migrates ventrolaterally through the anterior portion of each sclerotome. Sclerotomes are blocks of mesodermal cells, derived from somites, that will differentiate into the vertebral cartilage of the spine. The migration of the trunk neural crest initiates in a crowded environment around the posterior neural tube, where the cells are in close contact with the prospective epidermis and paraxial mesoderm. This restricts the migration of trunk neural crest cells to specific predetermined routes and makes them susceptible to the physical and molecular cues afforded by the surrounding environment.
The path taken by the migrating trunk neural crest cells is controlled by the extracellular matrices surrounding the neural tube. One set of proteins promotes migration. These proteins include fibronectin, laminin, tenascin, various collagen molecules, and proteoglycans, and they are seen throughout the matrix encountered by the neural crest cells. Another set of proteins impedes migration and provides the specificity for cellular movements. The main proteins involved in this restriction of neural crest cell migration are the ephrin proteins. These proteins are expressed in the posterior section of each sclerotome, and wherever they are, neural crest cells do not go.
The trunk neural crest cells that remain in the sclerotome form the dorsal root ganglia containing the sensory neurons. Those cells that continue more ventrally form the sympathetic ganglia, the adrenal medulla, and the nerve clusters surrounding the aorta. In birds and mammals, these cells migrate ventrally through the anterior but not through the posterior section of the sclerotomes.
Plymouth Rock Chicken Marketing: Targeting the Right Niche
You may want to see also
Frequently asked questions
The neural crest is a group of cells that delaminate from the dorsal neural tube and migrate throughout the developing embryo.
Neural crest migration occurs during embryogenesis. In the early embryo, trunk neural crest migration begins once the trunk neural tube has developed at least 7 somites.
Neural crest cells can take one of three major pathways. The first pathway involves cells migrating to the first pharyngeal arch, forming the jawbones and ear bones. The second pathway involves cells populating the second pharyngeal arch, forming the hyoid cartilage of the neck. The third pathway involves cells migrating through the rostral half of somites, differentiating into sensory and sympathetic neurons of the peripheral nervous system.
Neural crest cells migrate in loose clusters with a few single cells in the lead. They are elongated and aligned parallel to the direction of migration. They move through spaces between the basal laminae that define the basal surface of the epidermis and the developing dermomyotome.