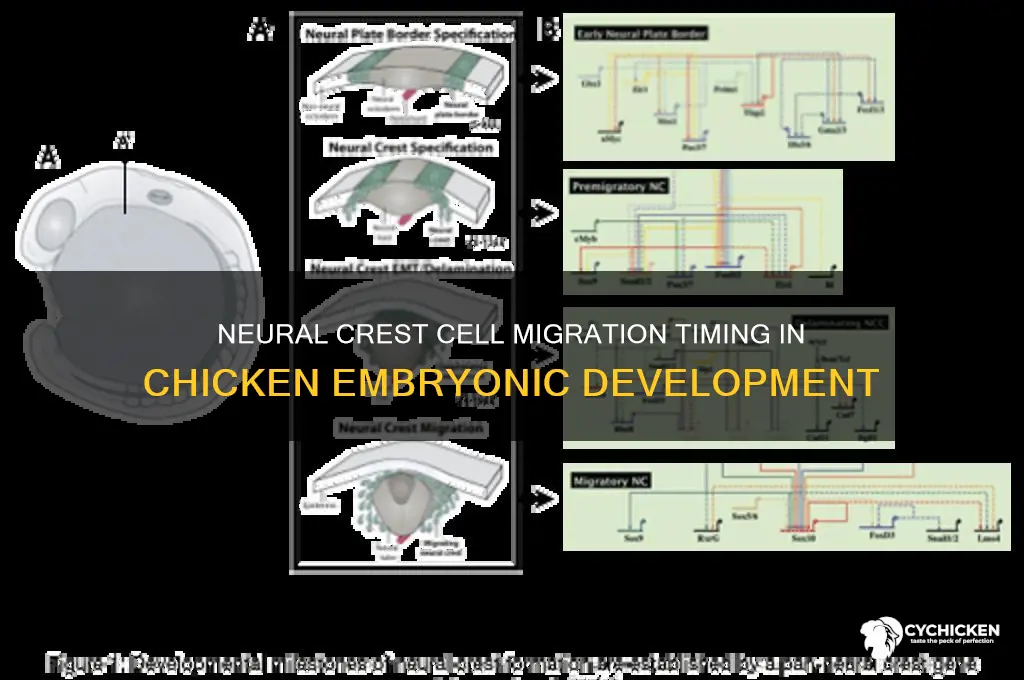
Neural crest cells, a transient and multipotent cell population, play a crucial role in vertebrate development, contributing to diverse structures such as the peripheral nervous system, craniofacial cartilage, and pigment cells. In chickens, a widely studied model organism, the migration of neural crest cells begins around embryonic stage HH4 to HH6 (Hamburger-Hamilton stages), approximately 18 to 24 hours after incubation. This process is initiated when neural crest cells delaminate from the dorsal region of the neural tube and embark on their journey to specific target locations throughout the embryo. The precise timing and regulation of this migration are tightly controlled by a complex interplay of signaling pathways, including Wnt, BMP, and FGF, ensuring proper differentiation and morphogenesis of neural crest-derived tissues. Understanding the onset and mechanisms of neural crest migration in chickens provides valuable insights into developmental biology and evolutionary processes.
| Characteristics | Values |
|---|---|
| Stage of Development | Hamburger-Hamilton (HH) Stage 8-9 |
| Time Post-Incubation | Approximately 18-24 hours |
| Location of Origin | Dorsal neural tube |
| Trigger for Migration | Induction by signals from the non-neural ectoderm (e.g., BMP, Wnt) |
| Direction of Migration | Ventrally and laterally through the rostral and caudal neuropores |
| Cell Behavior | Epithelial-to-mesenchymal transition (EMT) |
| Key Molecular Markers | Expression of Slug, Snail2, HNK-1, and Sox10 |
| Migratory Pathways | Dorsolateral pathways, stream into the pharyngeal arches and trunk |
| Environmental Cues | Guided by chemotactic signals (e.g., FGF, SDF-1) and extracellular matrix |
| Functional Significance | Formation of craniofacial structures, peripheral nervous system, etc. |
| Species-Specific Timing | Earlier than in mammals (e.g., mice start at E8.5) |
Explore related products






What You'll Learn
![]()
Neural crest induction timing in avian embryos
The induction of neural crest cells in avian embryos is orchestrated by a complex interplay of signaling pathways, including Wnt, BMP, FGF, and Notch. These pathways are active in the prospective neural crest region prior to migration, ensuring that cells acquire the necessary molecular identity to delaminate and embark on their migratory paths. Experimental studies have shown that perturbation of these signaling pathways during the critical window of HH stages 8 to 10 can disrupt neural crest induction and subsequent migration, highlighting the importance of precise timing in this process. For instance, inhibition of Wnt signaling during this period results in a failure of neural crest cells to form, while excessive BMP signaling can lead to an overproduction of neural crest cells.
Migration of neural crest cells in chickens begins shortly after their induction, typically around HH stage 10 to 12. At this stage, the cells undergo an epithelial-to-mesenchymal transition (EMT), losing their epithelial characteristics and gaining motility. This transition is marked by downregulation of epithelial markers such as E-cadherin and upregulation of mesenchymal markers like N-cadherin and fibronectin. The migratory pathways of neural crest cells are highly stereotyped, with distinct streams of cells moving ventrally, dorsolaterally, and into the branchial arches to contribute to various tissues and structures, including the peripheral nervous system, facial cartilages, and pigment cells.
The timing of neural crest migration is crucial for proper embryonic development, as delays or disruptions can lead to congenital abnormalities. For example, DiGeorge syndrome in humans, characterized by defects in craniofacial development and cardiac anomalies, is linked to aberrant neural crest migration. In avian embryos, the precise timing of migration ensures that neural crest cells reach their target destinations at the appropriate developmental stages, allowing for the coordinated formation of complex tissues and organs. Thus, understanding the temporal dynamics of neural crest induction and migration in chickens provides valuable insights into both normal development and developmental disorders.
In summary, neural crest induction in avian embryos occurs during the early stages of neural tube formation, specifically at HH stages 8 to 10, with migration commencing shortly thereafter. This process is regulated by a network of signaling pathways and is essential for the formation of multiple cell types and tissues. The precise timing of these events underscores their importance in ensuring proper embryonic development, making the chicken embryo an invaluable model for studying neural crest biology.
Chicken Worm Infestation: What Are the Risks?
You may want to see also
Explore related products






![]()
Molecular signals triggering migration initiation
The initiation of neural crest cell migration in chickens is a tightly regulated process orchestrated by a complex interplay of molecular signals. This process begins around Hamburger-Hamilton (HH) stage 9, coinciding with the closure of the neural tube. One of the earliest and most critical signals is the downregulation of BMP (Bone Morphogenetic Protein) signaling in the dorsal neural tube. BMPs, which are secreted by the non-neural ectoderm, repress neural crest cell formation and migration. As neural fold elevation progresses, the expression of Noggin and Chordin, BMP antagonists secreted by the dorsal neural tube, increases. These antagonists bind to BMPs, effectively reducing their activity and creating a permissive environment for neural crest cells to delaminate and migrate.
Simultaneously, Wnt signaling plays a pivotal role in triggering migration initiation. Wnt ligands, particularly Wnt1 and Wnt3a, are expressed in the dorsal neural tube and activate the canonical Wnt/β-catenin pathway. This activation upregulates the expression of transcription factors such as Slug and Snail2, which are essential for neural crest cell delamination and epithelial-to-mesenchymal transition (EMT). Slug and Snail2, in turn, downregulate cadherin expression, reducing cell-cell adhesion and enabling neural crest cells to detach from the neural tube and initiate migration.
Another critical signaling pathway is FGF (Fibroblast Growth Factor) signaling. FGFs, particularly FGF8, are expressed in the hindbrain and adjacent ectoderm, creating a gradient that guides neural crest cells toward their migratory paths. FGF signaling activates downstream effectors such as MAPK/ERK, which promote cell proliferation and migration. Additionally, FGFs synergize with Wnt signaling to enhance the expression of EMT-related genes, further facilitating the transition from a stationary to a migratory state.
Notch signaling also contributes to the initiation of neural crest cell migration. Notch ligands, such as Delta1, are expressed in the dorsal neural tube, and their interaction with Notch receptors on neural crest cells activates the Notch pathway. This activation leads to the upregulation of Hairy1 and Hairy2, transcription factors that promote neural crest cell delamination. Notch signaling works in concert with Wnt and FGF pathways to ensure a coordinated and timely initiation of migration.
Finally, Chemokine signaling provides directional cues for migrating neural crest cells. Chemokines like SDF-1 (Stromal Cell-Derived Factor 1) and their receptors, such as CXCR4, establish a chemotactic gradient that guides cells along their migratory routes. SDF-1 is expressed in target tissues, such as the branchial arches, and acts as a chemoattractant for CXCR4-expressing neural crest cells. This chemokine-mediated guidance ensures that neural crest cells migrate to their appropriate destinations, where they differentiate into diverse cell types.
In summary, the initiation of neural crest cell migration in chickens is triggered by a coordinated network of molecular signals, including BMP antagonists, Wnt, FGF, Notch, and chemokine signaling. These pathways work synergistically to regulate EMT, delamination, and directed migration, ensuring the successful dispersal of neural crest cells to their target tissues. Understanding these molecular mechanisms provides critical insights into the developmental processes underlying neural crest cell migration.
Chicken Leg Quarter: Weight and Nutrition Facts
You may want to see also
Explore related products






![]()
Role of epithelial-mesenchymal transition (EMT) in migration
Neural crest cells in chickens initiate migration during embryonic development, specifically around stages 8-10 of Hamburger-Hamilton (HH) development, which corresponds to approximately 18-24 hours after incubation. This process is crucial for the formation of various tissues and structures, including the peripheral nervous system, facial cartilages, and pigment cells. A key mechanism underlying the migration of neural crest cells is the epithelial-mesenchymal transition (EMT), a biological process that enables cells to transition from a stationary epithelial state to a motile mesenchymal state. EMT is characterized by the loss of cell-cell adhesion, reorganization of the cytoskeleton, and acquisition of migratory and invasive properties, all of which are essential for neural crest cell migration.
During EMT, neural crest cells undergo significant molecular changes that facilitate their detachment from the neural tube and subsequent migration. One of the hallmark features of EMT is the downregulation of epithelial markers, such as E-cadherin, which mediates cell-cell adhesion. This reduction in E-cadherin expression allows neural crest cells to dissociate from neighboring cells, a prerequisite for migration. Concurrently, there is an upregulation of mesenchymal markers, including N-cadherin, fibronectin, and vimentin, which promote cell motility and interaction with the extracellular matrix (ECM). These molecular changes are orchestrated by a network of transcription factors, including Snail, Slug, Twist, and ZEB, which are master regulators of EMT.
The role of EMT in neural crest cell migration is further highlighted by the involvement of specific signaling pathways. For instance, the Wnt, FGF, and BMP pathways play critical roles in inducing EMT by activating the aforementioned transcription factors. These pathways are temporally and spatially regulated to ensure that EMT occurs at the appropriate developmental stage and location. Additionally, the Notch signaling pathway has been implicated in fine-tuning EMT, ensuring that neural crest cells undergo a partial EMT, which allows them to maintain some epithelial characteristics while acquiring mesenchymal traits necessary for migration.
Cytoskeletal reorganization is another critical aspect of EMT that enables neural crest cell migration. During EMT, the actin cytoskeleton undergoes dynamic changes, leading to the formation of filopodia and lamellipodia, which are cellular protrusions essential for directed migration. Rho GTPases, such as RhoA, Rac1, and Cdc42, regulate these cytoskeletal changes by controlling actin polymerization and myosin contractility. Furthermore, the interaction between neural crest cells and the ECM is mediated by integrins, which are upregulated during EMT and facilitate cell adhesion, migration, and signaling.
In the context of chicken neural crest cell migration, EMT is not only a cellular process but also a spatially organized event. Neural crest cells delaminate from specific regions of the neural tube, known as the dorsal neural tube, where EMT is induced by signals from the surrounding tissues, including the ectoderm and paraxial mesoderm. This regional specificity ensures that neural crest cells migrate along defined pathways to reach their target destinations. Disruption of EMT, either genetically or experimentally, leads to defects in neural crest cell migration, resulting in craniofacial abnormalities and other developmental anomalies, underscoring the critical role of EMT in this process.
Understanding the role of EMT in neural crest cell migration has broader implications for developmental biology and regenerative medicine. The molecular mechanisms governing EMT in neural crest cells provide insights into similar processes in cancer metastasis, where EMT enables tumor cells to invade and migrate to distant sites. By studying EMT in the context of neural crest cell migration, researchers can identify potential therapeutic targets to modulate EMT in pathological conditions while ensuring proper embryonic development. Thus, the EMT process in neural crest cells serves as a paradigm for understanding cell migration in both physiological and disease contexts.
Local Chicken Farmers: Portland's Best Kept Secret
You may want to see also
Explore related products






![]()
Spatial patterns of neural crest migration pathways
Neural crest cells (NCCs) are a transient, multipotent cell population that arises from the dorsal neural tube and migrate extensively to contribute to diverse tissues in vertebrates. In chickens, the migration of neural crest cells begins around Hamburger-Hamilton (HH) stage 8 to 9, which corresponds to approximately 18 to 24 hours of incubation. This timing is critical, as it marks the initiation of their journey from the neural tube to their target destinations throughout the embryo. Understanding the spatial patterns of neural crest migration pathways is essential for deciphering their roles in development and the mechanisms guiding their movement.
The migration of NCCs in chickens follows distinct pathways, which are spatially organized along the rostrocaudal (head-to-tail) and dorsoventral (back-to-belly) axes. Cranial NCCs, originating from the anterior neural tube, migrate ventrally and laterally to form structures such as the craniofacial skeleton, connective tissues, and sensory ganglia. These cells follow well-defined streams, often guided by extracellular matrix components and chemotactic signals. For instance, the mandibular and hyoid arches receive NCCs via specific pathways that ensure proper patterning of the facial region. The spatial precision of these pathways is crucial for the correct assembly of craniofacial structures.
Trunk NCCs, arising from the posterior neural tube, exhibit different migration patterns. These cells migrate ventrally through the somites, forming the dorsal root ganglia and contributing to the sympathetic nervous system. Additionally, a subset of trunk NCCs migrates dorsolaterally to form the pigment cells of the skin. The spatial organization of these pathways is tightly regulated by interactions with surrounding tissues, such as the somites and ectoderm, which provide both permissive and instructive cues. Disruptions in these pathways can lead to congenital defects, highlighting their developmental significance.
Cardiac NCCs represent another distinct population, migrating from the posterior region of the neural tube to contribute to the outflow tract of the heart. Their pathway is spatially confined, ensuring precise integration into the developing cardiovascular system. This migration is guided by signals from the pharynx and adjacent mesoderm, underscoring the importance of tissue interactions in establishing spatial patterns. Aberrations in cardiac NCC migration are associated with congenital heart defects, emphasizing the clinical relevance of understanding these pathways.
In summary, the spatial patterns of neural crest migration pathways in chickens are highly organized and tailored to the specific functions of NCCs. Cranial, trunk, and cardiac NCCs follow distinct routes, guided by a combination of intrinsic programs and extrinsic signals. These pathways are established early in development, beginning around HH stage 8 to 9, and are essential for the formation of diverse tissues and organs. Investigating these spatial patterns not only provides insights into embryonic development but also informs our understanding of human congenital disorders linked to neural crest defects.
McDonald's Hot 'n Spicy Chicken Sandwich: Nationwide Availability?
You may want to see also
Explore related products






![]()
Environmental factors influencing migration onset in chickens
Neural crest cells (NCCs) in chickens initiate migration during embryonic development, typically around stages 8-10 (approximately 18-24 hours post-fertilization). This process is tightly regulated by both genetic and environmental factors. Environmental influences can significantly modulate the onset and efficiency of NCC migration, impacting subsequent tissue formation and embryonic health. Understanding these factors is crucial for developmental biology research and poultry science, as disruptions can lead to congenital defects or reduced hatchability.
Temperature is a critical environmental factor affecting NCC migration in chickens. Embryonic development is highly sensitive to thermal conditions, with optimal temperatures ranging between 37.5°C and 38.5°C. Deviations from this range can delay or accelerate NCC migration. For instance, exposure to lower temperatures (e.g., 35°C) has been shown to slow down cellular processes, including NCC delamination and migration, due to reduced metabolic activity. Conversely, higher temperatures (e.g., 40°C) can induce cellular stress, impairing the cytoskeletal dynamics essential for NCC movement. Precise temperature control during incubation is therefore vital to ensure timely and coordinated NCC migration.
Oxygen levels also play a significant role in modulating NCC migration onset. Chicken embryos develop in a naturally hypoxic environment, with oxygen levels around 15-20% in the incubator. Hypoxia-inducible factors (HIFs) are activated under these conditions, promoting the expression of genes involved in NCC migration, such as *Snail2* and *Twist*. However, excessive hypoxia (e.g., <10% O2) or hyperoxia (e.g., >40% O2) can disrupt this balance. Hyperoxia, for example, increases oxidative stress, damaging cellular structures and impairing NCC motility. Researchers must carefully regulate oxygen levels to mimic the physiological environment and support normal NCC migration.
Mechanical cues from the embryo's surroundings can further influence NCC migration. The stiffness of the extracellular matrix (ECM) and the physical forces exerted during incubation affect cellular behavior. Studies have shown that alterations in ECM stiffness, achieved through experimental manipulation, can either enhance or inhibit NCC migration. Additionally, vibrations or physical disturbances during incubation may disrupt the coordinated movement of NCCs, leading to aberrant migration patterns. Maintaining a stable and controlled incubation environment minimizes mechanical perturbations, ensuring proper NCC migration.
Chemical exposure during early embryonic development can have profound effects on NCC migration. Environmental toxins, such as heavy metals or pesticides, can interfere with signaling pathways critical for NCC delamination and migration. For example, exposure to cadmium has been linked to reduced expression of *HNK-1*, a surface marker essential for NCC migration. Similarly, endocrine-disrupting chemicals (EDCs) can alter hormone levels, indirectly affecting NCC behavior. Rigorous screening and regulation of chemical contaminants in poultry farming environments are essential to prevent developmental abnormalities associated with NCC migration defects.
In summary, environmental factors such as temperature, oxygen levels, mechanical cues, and chemical exposure significantly influence the onset and progression of NCC migration in chickens. Precise control of these factors during incubation is critical to ensure optimal embryonic development and hatchability. Future research should continue to explore the interplay between environmental conditions and NCC migration, providing insights for both developmental biology and poultry production practices.
Ground Chicken vs Turkey: Which Has More Fat?
You may want to see also
Frequently asked questions
Neural crest cell migration in chicken embryos typically starts around Hamburger-Hamilton (HH) stage 8 to 9, which corresponds to approximately 14 to 18 hours after incubation.
The initiation of neural crest cell migration is triggered by a combination of signaling pathways, including BMP, Wnt, and FGF, which regulate the epithelial-to-mesenchymal transition (EMT) and induce the expression of migration-related genes.
No, neural crest cell migration is spatially and temporally regulated. Cells from different axial levels (e.g., cranial, trunk, vagal) begin migration at slightly different times, with cranial neural crest cells migrating first, followed by trunk and vagal populations.
The neural tube provides essential signals and structural support for neural crest cell migration. As the neural tube closes, it releases signals that guide neural crest cells to delaminate and migrate along specific pathways to their target destinations.