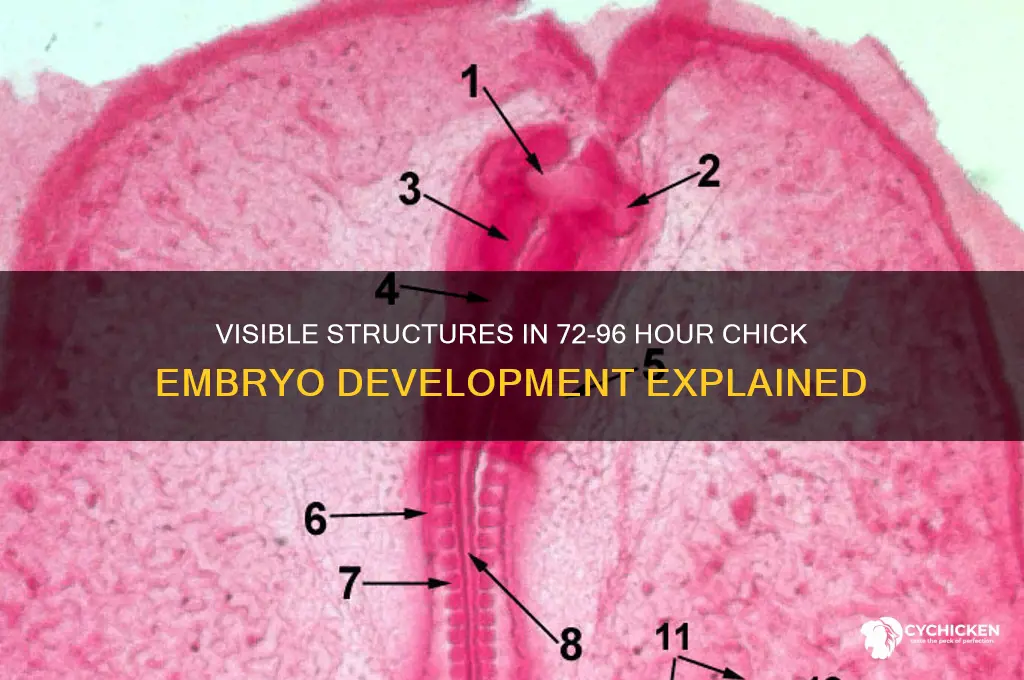
The 72-96 hour stage of chick embryo development, often referred to as the late gastrulation to early organogenesis period, is a critical phase marked by rapid morphological changes and the emergence of distinct structures. During this time, the embryo transitions from a three-layered structure (ectoderm, mesoderm, and endoderm) to a more complex organization, with visible formation of key features such as the neural tube, somites, and the primitive heart tube. The neural folds fuse to form the neural tube, which will give rise to the central nervous system, while paired somites appear along the paraxial mesoderm, laying the foundation for future vertebrae and skeletal muscle. Additionally, the primitive streak begins to regress, and the cardiovascular system initiates development, with the heart tube becoming visible and beginning to beat, establishing the early circulatory system. These structures are readily observable under a stereomicroscope, providing valuable insights into the early stages of vertebrate development.
Explore related products






What You'll Learn
- Yolk Sac Formation: Visible as a large, vascularized sac attached to the embryo
- Amnion Development: Thin membrane enclosing the embryo, filled with amniotic fluid
- Allantois Emergence: Outpouching from hindgut, aids in waste storage and gas exchange
- Somite Segmentation: Paired blocks along the neural tube, forming mesoderm
- Neural Tube Closure: Fusing to form the brain and spinal cord precursors
![]()
Yolk Sac Formation: Visible as a large, vascularized sac attached to the embryo
Between 72 and 96 hours of chick embryo development, one of the most prominent and critical structures visible is the Yolk Sac. At this stage, the yolk sac is clearly discernible as a large, vascularized sac that remains attached to the embryo. This structure is essential for the embryo’s early development, serving as a vital source of nutrients and playing a key role in the formation of the circulatory system. The yolk sac’s vascular network, known as the vitelline vasculature, becomes increasingly prominent during this period, facilitating the exchange of nutrients and waste products between the embryo and the yolk.
The formation of the yolk sac is a dynamic process that begins earlier in development but becomes more pronounced by 72–96 hours. Initially, the yolk sac appears as a simple membrane surrounding the yolk, but it rapidly develops a complex network of blood vessels. These vessels are extensions of the embryo’s developing circulatory system, connecting the yolk sac to the embryo via the vitelline vessels. Under a microscope or in high-resolution imaging, the yolk sac’s vascularization is evident as a dense, intricate pattern of blood vessels that radiate across its surface. This vascular network is crucial for the embryo’s growth, as it allows for the absorption of nutrients from the yolk, which are then transported to the developing tissues.
By 72–96 hours, the yolk sac is not only larger but also more distinct in its attachment to the embryo. It is typically positioned ventrally relative to the embryo, connected by the vitelline duct (ombilical yolk stalk). This attachment is critical for maintaining the nutrient supply and supporting the early stages of organogenesis. The yolk sac’s size and vascularization make it one of the most visually striking features of the embryo at this stage, often dominating the field of view in developmental studies. Its prominence allows researchers to easily identify and assess its development, making it a key structure for evaluating embryonic health and progress.
The vascularized yolk sac also plays a pivotal role in hematopoiesis, the formation of blood cells. During this developmental window, primitive blood cells begin to emerge within the yolk sac’s vasculature, marking the initial stages of the embryo’s blood system. This process is closely linked to the yolk sac’s nutrient absorption function, as the developing blood cells rely on the nutrients derived from the yolk. Observing the yolk sac’s vascularization and its connection to the embryo provides valuable insights into the coordinated development of the circulatory and digestive systems.
In summary, the yolk sac’s formation and vascularization between 72 and 96 hours of chick embryo development are critical processes that support the embryo’s growth and organogenesis. Visible as a large, vascularized sac attached to the embryo, it serves as a nutrient reservoir, a site for early blood cell formation, and a key component of the developing circulatory system. Its prominence and accessibility make it an essential structure for studying early embryonic development and assessing the health of the growing organism.
Who's the Colonel in KFC's Chicken and Waffles Ad?
You may want to see also
Explore related products






![]()
Amnion Development: Thin membrane enclosing the embryo, filled with amniotic fluid
At 72 to 96 hours of chick embryo development, the amnion, a thin, translucent membrane, becomes clearly visible and functional. This structure is crucial for the embryo's protection and development, as it encloses the embryo and is filled with amniotic fluid. The amnion initially forms as a simple layer of cells derived from the epiblast, one of the primary cell layers in the early embryo. Over the course of these 24 to 48 hours, it expands and differentiates, creating a distinct sac that surrounds the developing embryo. This process is essential for establishing the amniotic cavity, which provides a buffered environment for growth.
The amniotic fluid within the amnion plays a vital role in supporting the embryo. It acts as a cushion, protecting the embryo from mechanical shocks and allowing for free movement, which is critical for proper musculoskeletal development. Additionally, the fluid helps regulate temperature and prevents the tissues from sticking together, ensuring that the embryo can develop without deformities. By 72 to 96 hours, the amniotic fluid is visibly present, and its volume increases as the embryo grows, maintaining a stable environment for further organogenesis.
During this stage, the amnion also begins to interact with other extraembryonic membranes, such as the yolk sac and allantois. The yolk sac, which provides nutrients, remains separate from the amnion but is closely associated with it. The allantois, responsible for waste storage and gas exchange, starts to expand and may come into contact with the amnion, though they remain distinct structures. These interactions highlight the coordinated development of extraembryonic tissues to support the embryo.
Microscopic examination at 72 to 96 hours reveals the amnion as a single layer of flattened epithelial cells, known as amnioblasts, supported by a thin basement membrane. These cells secrete the amniotic fluid and maintain the integrity of the membrane. The simplicity of this structure is deceptive, as it performs multiple functions, including protection, fluid regulation, and support for embryonic movements. The clarity of the amnion at this stage makes it a key structure for observational studies in developmental biology.
In summary, between 72 and 96 hours of chick embryo development, the amnion undergoes significant growth and differentiation, becoming a visible and functional structure. Its role in enclosing the embryo and maintaining amniotic fluid is critical for protection, movement, and overall development. The interaction with other extraembryonic membranes and its distinct cellular composition further underscore its importance in this critical window of embryogenesis. Observing the amnion at this stage provides valuable insights into the early stages of avian development.
Chicken Steak: Myth or Menu Reality? Exploring the Culinary Truth
You may want to see also
Explore related products






![]()
Allantois Emergence: Outpouching from hindgut, aids in waste storage and gas exchange
During the critical developmental window of 72-96 hours in a chick embryo, the allantois begins to emerge as a distinct structure. This emergence is characterized by the formation of an outpouching from the hindgut, marking a significant milestone in embryonic development. The allantois initially appears as a small, fluid-filled sac that extends posteriorly from the hindgut region. This outpouching is a result of localized growth and morphogenesis, driven by genetic and molecular signals that coordinate tissue differentiation. Understanding this process is essential, as the allantois plays a pivotal role in supporting the embryo's metabolic needs during its early stages.
The primary functions of the allantois include waste storage and gas exchange, both of which are critical for the embryo's survival. As the embryo grows, metabolic waste products accumulate, and the allantois serves as a temporary storage site for these wastes, preventing their buildup in the embryonic environment. Simultaneously, the allantois facilitates gas exchange by increasing the surface area available for diffusion of oxygen and carbon dioxide. This is particularly important in avian embryos, where the eggshell limits direct gas exchange with the external environment. The allantois expands and eventually fuses with the chorion, forming the chorioallantoic membrane (CAM), which becomes the primary site for respiratory gas exchange.
The emergence of the allantois is closely coordinated with other developmental processes occurring at 72-96 hours. During this period, the embryo undergoes rapid cell division and differentiation, and the allantois development is integrated with the formation of the amnion, yolk sac, and other extraembryonic membranes. The hindgut, from which the allantois originates, is part of the developing digestive system, and its outpouching reflects the embryo's increasing complexity and specialization. Observing the allantois at this stage provides valuable insights into the spatial and temporal dynamics of embryonic organogenesis.
Microscopic examination of the chick embryo at 72-96 hours reveals the allantois as a transparent, fluid-filled structure adjacent to the hindgut. Its growth is evident as it extends posteriorly, often appearing as a distinct bulge in the embryonic tissue. This visibility makes the allantois a key structure to identify during developmental studies, allowing researchers to assess the progress of embryonic growth and the health of the embryo. The clarity of its emergence underscores the precision of developmental biology, where each structure forms at a specific time and location to fulfill its designated function.
In summary, the emergence of the allantois as an outpouching from the hindgut between 72-96 hours in the chick embryo is a critical event in early development. Its formation supports waste storage and gas exchange, addressing the metabolic demands of the growing embryo. By studying this process, researchers gain a deeper understanding of the intricate coordination required for successful embryonic development. The allantois, with its distinct morphology and function, stands as a testament to the complexity and elegance of developmental biology.
Weight Watchers: Chicken & Dumplings Point Value
You may want to see also
Explore related products






![]()
Somite Segmentation: Paired blocks along the neural tube, forming mesoderm
Between 72 and 96 hours of development, the chick embryo undergoes significant morphological changes, and one of the most prominent features visible during this stage is somite segmentation. Somites are paired, metameric blocks of mesoderm that form along either side of the neural tube. This process is a critical step in the development of the vertebrate body plan, as somites give rise to essential structures such as the vertebral column, ribs, dermis, and skeletal muscle. Observing the chick embryo at this stage reveals a distinct, segmented appearance along the anterior-posterior axis, with somites arranged in a rostro-caudal sequence.
The formation of somites begins with the paraxial mesoderm, a region of mesoderm adjacent to the neural tube. As the embryo develops, this mesoderm undergoes a process called somitogenesis, where it condenses and segments into individual somites. Each somite is initially connected to the neural tube and the notochord, a rod-like structure that provides structural support during early development. The segmentation of somites is highly regulated by molecular signals, including the oscillatory expression of genes in the segmentation clock, which ensures the precise and repetitive formation of these blocks.
Under a microscope or in high-resolution images, the somites appear as distinct, bilaterally symmetrical blocks flanking the neural tube. Their shape is initially epithelial, with a clear boundary separating each somite from its neighbors. The anterior and posterior boundaries of each somite are particularly well-defined, reflecting the coordinated action of signaling pathways such as Wnt, FGF, and Notch. These boundaries are crucial for the proper differentiation and patterning of the tissues derived from the somites.
As development progresses within this 72-96 hour window, the somites begin to differentiate into three main regions: the sclerotome, dermatome, and myotome. The sclerotome gives rise to the vertebral column and ribs, the dermatome contributes to the dermis of the skin, and the myotome forms skeletal muscle. This regionalization is evident in the changing morphology of the somites, with cells migrating and reorganizing to form these distinct layers. The sclerotome cells, for example, can be observed migrating towards the notochord and neural tube, where they will eventually form the cartilaginous templates of the vertebrae.
In summary, somite segmentation is a hallmark of the 72-96 hour chick embryo, characterized by the formation of paired mesodermal blocks along the neural tube. This process is not only visually striking but also functionally vital, as it lays the foundation for the musculoskeletal system. By examining the chick embryo at this stage, researchers can gain insights into the molecular and cellular mechanisms driving somitogenesis, a process conserved across vertebrates. The clear segmentation and subsequent differentiation of somites make this developmental window an ideal period for studying early embryonic patterning and tissue specification.
Why Bone-In Chicken Makes My Stomach Turn: Unraveling the Discomfort
You may want to see also
Explore related products




![]()
Neural Tube Closure: Fusing to form the brain and spinal cord precursors
Between 72 and 96 hours of development, the chick embryo undergoes a critical process known as neural tube closure, which is essential for the formation of the brain and spinal cord precursors. At this stage, the neural plate, a thickened ectodermal region along the midline of the embryo, begins to fold and fuse, transforming into the neural tube. This process is highly coordinated and involves the precise migration and differentiation of cells. By 72 hours, the neural folds, which are the elevated edges of the neural plate, start to elevate and converge toward the midline. This convergence is driven by molecular signals, including morphogens like Sonic Hedgehog (Shh) and Bone Morphogenetic Proteins (BMPs), which regulate cell behavior and ensure proper folding.
As the neural folds continue to elevate, they gradually approach each other, forming a groove known as the neural groove. By 84–96 hours, the neural folds fuse at the midline, converting the neural groove into a closed neural tube. This closure occurs in a rostral-to-caudal direction, meaning it begins near the future brain region and progresses toward the spinal cord region. The successful fusion of the neural folds is crucial, as any disruption can lead to neural tube defects, such as spina bifida or anencephaly. During this period, the anterior portion of the neural tube, which will give rise to the brain, begins to expand and differentiate into distinct regions known as primary brain vesicles.
Microscopic examination of the chick embryo at this stage reveals the neural tube as a distinct, hollow structure running along the dorsal side of the embryo. The closure of the neural tube is accompanied by the formation of other visible structures, such as the somites, which are paired blocks of mesoderm flanking the neural tube and will develop into vertebral and skeletal muscle tissues. The notochord, a rod-like structure located beneath the neural tube, also becomes more defined and plays a key role in patterning the overlying neural tissue.
The molecular mechanisms underlying neural tube closure involve a complex interplay of genes and signaling pathways. For example, the planar cell polarity (PCP) pathway regulates cell shape changes and coordinated movement of the neural folds, while the Wnt signaling pathway influences cell proliferation and differentiation. Additionally, extracellular matrix components, such as laminin and fibronectin, provide structural support and facilitate cell migration during closure. These processes ensure that the neural tube forms a seamless structure, which is essential for the subsequent development of the central nervous system.
By 96 hours, the neural tube is fully closed, and the embryo enters the next phase of neural development, where the tube begins to regionalize into distinct domains. The anterior portion starts to balloon into three primary brain vesicles: the prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain). Simultaneously, the posterior neural tube forms the spinal cord precursor. This regionalization is guided by gradients of signaling molecules, which establish the body plan and lay the foundation for the complex neural circuitry that will emerge in later developmental stages.
In summary, neural tube closure between 72 and 96 hours in the chick embryo is a pivotal event in the formation of the brain and spinal cord precursors. This process involves the elevation, convergence, and fusion of the neural folds, driven by precise molecular and cellular mechanisms. The successful closure of the neural tube is essential for proper neural development, and its study provides valuable insights into the early stages of embryogenesis and the origins of neural tube defects. Observing these structures during this critical window offers a unique opportunity to understand the foundational steps of nervous system formation.
Gluten-Free Parmesan Crusted Chicken at Noodles & Company
You may want to see also
Frequently asked questions
At 72-96 hours, visible structures include the amniotic membrane, yolk sac, allantois, and the developing embryo with distinct head, tail, and somites (early segments of the body).
Yes, the heart is visible and begins to beat during this stage, often seen as a pulsating structure in the anterior region of the embryo.
Limb buds start to form during this period, appearing as small swellings on the lateral sides of the embryo, marking the beginnings of wings and legs.
The yolk sac provides nutrients and is connected to the embryo via the vitelline membrane, supporting early development before the digestive system is functional.
The allantois begins to expand during this stage, serving as a respiratory organ for gas exchange and later contributing to the formation of the urinary bladder.










![Role Models [Blu-ray]](https://m.media-amazon.com/images/I/81wJlwDSY+L._AC_UL320_.jpg)




















